養殖研究所田中秀樹研發組長專訪
台灣鰻訊 第237期 2004-03-12 林雪貞譯自2004.01.10養殖新聞
在2003年的業界,最大的話題人物應該就是三重縣的獨立行政法人水產綜合研究中心養殖研究所的田中秀樹組長。研究所成功進行了鰻魚苗的人工生產,為鰻魚的完全養殖工作打開了大門,這對負面消息頻傳的鰻魚業界而言,可是久違了的好消息。業界相關人士頻繁的造訪養殖研究所,並不斷有來自各地的演講邀請,人氣是發燒又發熱。在本篇報導中,我們一面介紹田中專家這位發燒人物的專訪,並且歸納與整理鰻魚的完全養殖化的課題及展望。
養殖研究所位在三重縣伊勢市往南車程約一小時的地方。今年有許多鰻魚業界的各個團體造訪養殖研究所。從大隅地區養鰻漁業協會、一色鰻魚漁業協會、到波崎共榮漁業協會等,以鰻魚苗為生命線的鰻魚養殖相關的許多業界團體為了要親眼目睹鰻魚苗的人工生產,而來到養殖研究所,並與田中專家進行了意見交換。此外,鰻魚資源保護對策協議會、東京鰻蒲燒商公會也分別邀請了田中專家前往東京發表演講等,業界對此的關心程度可見一斑。
我們請教了「熱門人物」田中秀樹專家在養殖研究所工作的心路歷程,以及鰻魚苗人工生產要導入商業使用上的各種課題。
田中專家首先談到了此次成功進行了鰻魚苗的人工生產。他表示「鰻魚苗要以人工生產出與捕撈的野生鰻魚相當的數量是困難的」。迄今為止,鰻魚苗的人工生產一直以來的課題就是魚苗的飼料。雖然鰻魚苗「有的也吃」一般幼魚所吃的飼料「輪蟲」,但魚苗「到底吃些什麼」一直不為人所知,因此向來研究的焦點就是飼料。而現在鰻魚葉形魚苗的飼料餵食方式不是「讓魚苗來吃飼料」,而是「使魚苗碰觸到飼料便數順口把飼料吃進去」的情形。如照片中所示,魚苗雖有獨特像牙齒一般的構造,但「這只有將飼料咬入口中的作用,並沒有咀嚼的功能」。因此,「利用鰻魚本身夜行性的特性,當房間明亮時,鰻魚會往暗處移動的習性,在水槽底部擺滿飼料,將房間照明打亮,來使鰻魚往有飼料的底部移動碰觸之」。但是「除了鯊魚卵之外,鰻魚即使碰觸到其他的飼料也不會吃」。而不吃的原因之今尚不明瞭。
一般認為「魚類的幼魚在胃部尚為成長之前是不會積極覓食的」,這在鰻魚身上極為顯著。田中專家也對此感受良多,「不知道魚苗到底吃些什麼」「魚苗真是離世又脫俗」。田中專家表示「迄今為止我們嘗試過生物飼料、調配飼料、營養加強飼料等各種飼料」,其中「鯊魚卵是魚苗吃最多的」。而這個發現帶來了此次人工生產的成功。由於曾有研究報告指出在比目魚的初期飼料中,胜肽及磷蝦具有效用。我們使用了不二製油公司的胜肽及磷蝦萃取液。在1999年成功的讓鰻魚葉形前期魚苗發育成葉形魚苗,成功飼養到第256天。然而這個飼料無法接續下去,因此也不能作為決定性的飼料。由於胜肽中含有妨礙鈣質攝取的成分,因此將飼料做了進一步的改良,將減低了植酸(phytic acid)的大豆peptides及磷蝦分解物(日本水產)餵食給葉形魚苗,而成功的讓葉形魚苗發育成鰻魚苗。
田中專家表示「第一次看到發育成鰻魚苗時很是驚訝」。但由於當初「尚未達到公開發表的階段」,並需要進一步對後續狀況做確認,以及等待魚苗繼續發育成中間種苗後才做了公開發表。但田中專家表示「還有很多的課題」。其中之一就是「為期較長的幼魚期」。他指出「就像加吶魚及龍蝦等一樣,人工孵化及培育的時間比起天然野生的有較長的傾向,而且其個別差異也很大」。一般認為「野生魚可能一整天都在覓食」。此外還有成品率偏低的問題。「在受精率30%、孵化率10%中,能培育出的鰻魚苗比率僅有零點零幾的百分比」。而「後續的鰻魚苗及中間魚的培育生長尚未穩定」,因此要如何供應「日本鰻魚養殖所需的30噸,共約1億5千萬尾的鰻魚苗」將成為今後的課題。而「目前魚苗培育的成本相當高」,要發展到能商業使用「還需相當的時間」。田中專家並表示「『培育鰻魚苗』的夢想已經實現。接下來要讓『培育養殖用的種苗』這個夢想實現」,「現在一個循環約需1年。我打算以10年為目標來努力」,充分顯現出專家積極的意願。
而在業界相關人士中流傳的「鰻魚會在日本近海產卵」或是「在台灣及日本的近海都有鰻魚苗」的看法也傳到了田中專家的耳中。對此,田中專家表示「我請他們『讓我看看實際的魚苗』,但實際上卻沒有人拿來。這個說法不夠實際」。此外,以東大海洋研究所為主,一直進行著特定出鰻魚產卵地點的工作。其中一般推測鰻魚是在菲律賓近海400公尺的深海處產卵。對此田中專家表示「迄今為止的研究當中,低於19度以下時魚卵是不會孵化的,即使加壓也沒有作用。因此鰻魚產卵的地方應該是在水深200公尺以內的地方」,他對鰻魚產卵提出了個人研究經驗中所得出的見解。劃時代的研究成果,使得鰻魚業界引頸盼望的鰻魚完全養殖化更接近了一步,造成了業界的轟動。
吊書袋 (四)
但在鰻魚生殖生理機制尚未完全解析明白之前,人工催熟催生有其實用之價值。最近日本學者嘗試改良初期餌料系統,由傳統之無脊椎動物受精卵,輪蟲等之微生物,改為經過部份酵素消化或萃取之水溶性有機物質 (見去年之 養魚世界)。已可將鰻苗存活期延長至250天 (圖一)。事實上,此一策略並非憑空而來,乃是由野外調查所得之結果並印證其他學者理論而來。之一、由電子顯微鏡觀察而得之直接證據,以掃描式電子顯微鏡觀察野生柳葉幼苗之消化道內含物,發現生物之碎屑、糞粒、及浮游生物之膠質體殼,因此學者提出柳葉鰻並無特定之食性且可能以經分解的有機物為能量來源;之二、證據顯示,柳葉型魚苗的消化道及體表細胞具胞飲作用,能吸收溶解性大分子;之三、由免疫組織化學染色的證據顯示,柳葉鰻消化道中蛋白脢 (trypsin) 及几丁脢 (chitinase) 的活性很低,不僅如此,消化腺胰臟中,體制素 (somatosatin) 量很高而胰島素 (insulin) 及昇醣激素 (glucagon) 之量很低,顯示代謝調節激素的作用受抑制。
故由不同之學理基礎研究結果和日本學者之成功經驗,可以預測柳葉鰻苗之餌料攝取,也可由上皮細胞直接吸收大分子。至於是那一類之大分子,或許我們可以從其他魚種得到啟示。 例如Albula sp. (北梭魚, 其幼苗也需經柳葉體型之變態),在人工海水中經過十天的飢餓,體成份的變化,體脂肪及灰份減少50%、醣類及水份減少 80%,可是溶解性蛋白質維持不變。其他的研究證據指出,柳葉幼苗變態所需之能量也來自構撐扁平體型之膠狀物質 (glycosaminoglycan),此種物質為一種酸性黏多醣類 (acid mucopolysaccarides)。而浮游期愈長對外營性營養的依賴愈大。最近的調查報告指出,柳葉魚苗的組織代謝率,隨著體型變大而降低,而且大部份的耗能是用於將體膠狀物質 (glycosaminoglycan) 代謝以產生新的能量。是故在柳葉鰻之消化道內發現之尾蟲類膠質體殼,可能有其生理意義, 需進一步分析研究。仔魚胚胎發育的營養來自母魚,借由卵黃,在卵黃囊吸收完畢之後,幼魚必需自營生活。以人工孵化之鰻魚及野生鰻之生活史為例,卵黃囊約在一周至兩週內左右吸收完畢。而大洋浮游期可能至少長達數月至數年,假設柳葉鰻苗能由外界吸收營養物質,那麼早期柳葉鰻苗體成份之變化,應可以反映柳葉鰻苗由外界吸收營養物質的質與量,因為後其柳葉鰻苗不會消耗太多之能量來進行新陳代謝。這一點推論可由體制素,胰島素及昇醣激素的相對量得到支持。也可能預測柳葉體型的程度 (能量物質的堆積),可能對存活率相當重要。人工養殖就是要突破這一點天然限制,以外來的營養物質,代替先天之不足,但原則是:將鰻苗 '泡' 在 '蜜與牛奶' 中…。
主要參考文獻:
田中秀樹 (1999) 養殖研 News, 43:52-53.
Bishop, R. E. and Torres, J. J. (1999) Leptocephalus energetics: metabolism and excretion. J. Exp. Biol., 202(18):2485-2493.
Kruse,C., Strehlow,B., Schmidt,H., Muller,P.K. (1996) Presence of trypsin in distinctive body segments of leptocephalus larvare of Anguilliformes. Aquaculture, 142, 237-244.
L'Hermite,A., Ferrand,R., Dibois,M.P., Andersen,A.C. (1985) Detection of endocrine cells by immunofluorescence method in the gastroenteropancreatic system of the adult eel, glass-eel, and leptocephalic larva (Anguilla anguilla L.). Gen.Comp.Endocrinol., 58, 347-259.
Mochioka, N., Iwamizu, M. (1996) Diet of anguilloid larvae: leptocephali feed selectively on larvacean houses and fecal pellets. Mar. Biol., 125: 447-452.
Otake,T. (1996) Fine structure and function of the alimentary canal in leptocephali of the Japanese eel Anguilla japonica. Fish.Sci., 62, 28-34.
Pfeiler,E. (1991) Glycosaminoglycan composition of anguilliform and elopiform leptocephali. J.Fish Biol., 38, 533-540.
Pfeiler,E. (1999) Developmental physiology of elopomorph leptocephali. Comp.Biochem.Physiol., 123A, 113-128.
Tuesday, February 28, 2006
Sunday, February 26, 2006
News and Views
Nature 439, 926 (23 February 2006) | doi:10.1038/439926a
Spawning spot
Rosalind Cotter
The enigma of the eel life-cycle has become a little less enigmatic — at least as far as the Japanese eel, Anguilla japonica, is concerned. As described elsewhere in this issue (Nature 439, 929; 2006), Katsumi Tsukamoto has filled in a missing part of the picture of the species' ocean biology.
When they are mature, eels migrate from fresh water out into the open ocean, travelling thousands of kilometres to spawn. But the when and where of the actual spawning event, although suspected in general terms, have been hard to pin down. This is partly because of the technical challenges of isolating and identifying the tiny, transparent larvae just after they hatch. Tsukamoto has tackled those challenges to pinpoint the timing — which aptly coincided with a new moon in June — and strategic location of A. japonica spawning.

OCEAN RES. INST., UNIV. TOKYO
This fishing feat also meant that the newly hatched larvae could be photographed. As shown in this picture, teeth, jaws and pigmented eyes are evident within five days of hatching, when the larvae are still less than 5 millimetres long.
Thanks to the efforts in the 1920s of a Danish oceanographer, Johannes Schmidt, the spawning area of the European (A. anguilla) and American (A. rostrata) eels was identified as the Sargasso Sea in the western North Atlantic, spreading over some 5 million square kilometres. But it seems that the nursery of the Japanese eel, as identified by Tsukamoto, is much more compact — spawning is confined to a seamount northwest of Guam in the Philippine Sea, a location that enables the larvae to ride the northbound Kuroshio Current to their freshwater destinations in eastern Asia.
This discovery will be applauded by researchers specializing in eel biology, as well as by those intrigued by a life history that constitutes one of nature's wonders. But there is a commercial aspect too: eels are of great economic value, not least in Asia, and there is a pressing need for management and conservation of natural populations.
Nature 439, 926 (23 February 2006) | doi:10.1038/439926a
Spawning spot
Rosalind Cotter
The enigma of the eel life-cycle has become a little less enigmatic — at least as far as the Japanese eel, Anguilla japonica, is concerned. As described elsewhere in this issue (Nature 439, 929; 2006), Katsumi Tsukamoto has filled in a missing part of the picture of the species' ocean biology.
When they are mature, eels migrate from fresh water out into the open ocean, travelling thousands of kilometres to spawn. But the when and where of the actual spawning event, although suspected in general terms, have been hard to pin down. This is partly because of the technical challenges of isolating and identifying the tiny, transparent larvae just after they hatch. Tsukamoto has tackled those challenges to pinpoint the timing — which aptly coincided with a new moon in June — and strategic location of A. japonica spawning.

OCEAN RES. INST., UNIV. TOKYO
This fishing feat also meant that the newly hatched larvae could be photographed. As shown in this picture, teeth, jaws and pigmented eyes are evident within five days of hatching, when the larvae are still less than 5 millimetres long.
Thanks to the efforts in the 1920s of a Danish oceanographer, Johannes Schmidt, the spawning area of the European (A. anguilla) and American (A. rostrata) eels was identified as the Sargasso Sea in the western North Atlantic, spreading over some 5 million square kilometres. But it seems that the nursery of the Japanese eel, as identified by Tsukamoto, is much more compact — spawning is confined to a seamount northwest of Guam in the Philippine Sea, a location that enables the larvae to ride the northbound Kuroshio Current to their freshwater destinations in eastern Asia.
This discovery will be applauded by researchers specializing in eel biology, as well as by those intrigued by a life history that constitutes one of nature's wonders. But there is a commercial aspect too: eels are of great economic value, not least in Asia, and there is a pressing need for management and conservation of natural populations.
Friday, February 24, 2006
謎題有解 日本鰻產卵地 在關島海域
2006.02.24 中國時報
謎題有解 日本鰻產卵地 在關島海域
楊明暐/綜合報導
在日本料理中,美味的日本鰻魚一直是饕客心目中的頂饌珍饈。然而自古以來,日本鰻魚如何產卵一直是個未解之謎。日本科學家最近對關島附近海域的鰻魚苗進行基因比對之後,終於確認日本鰻魚原來是在該處海域繁衍後代。
日本鰻魚和牠們在大西洋的親戚一樣,在淡水中成長,然後再游回大海產卵。科學界早就知道生長在歐美淡水水域的鰻魚來自百慕達附近的馬尾藻海域,但沒人曉得日本鰻魚從哪裡來。
東京大學海洋研究所教授塚本勝巳領導的研究小組去年六月搭乘海洋研究船「白鳳丸」航行到馬里亞納群島西方靠近關島的海域。他們在海面下五百公尺深處撒下大型浮游生物網,並於「駿河海山」西方七十到一百公里處捕獲四百隻孵化約二至五天的鰻魚幼苗。經DNA鑑定,這些鰻苗確定是日本鰻。
研究人員按海流速度和孵化時間推算,認為位於北緯一四度、東經一四二度海底的「駿河海山」應該就是鰻魚產卵地。產下的鰻魚卵孵化後,隨北赤道海流漂向西方,而後再隨黑潮北上,經三個月到半年,游到台灣、中國大陸和日本海岸,鰻魚幼苗在這段期間會長大成為幼鰻,牠們接著便溯游至河流及湖泊中成長。
塚本將這項研究成果發表於廿三日出刊的最新一期英國《自然》周刊。他表示,確認鰻魚的產卵地之後,才能釐清鰻魚的迴游途徑,這項研究有助於保護日益稀少的日本鰻魚。
謎題有解 日本鰻產卵地 在關島海域
楊明暐/綜合報導
在日本料理中,美味的日本鰻魚一直是饕客心目中的頂饌珍饈。然而自古以來,日本鰻魚如何產卵一直是個未解之謎。日本科學家最近對關島附近海域的鰻魚苗進行基因比對之後,終於確認日本鰻魚原來是在該處海域繁衍後代。
日本鰻魚和牠們在大西洋的親戚一樣,在淡水中成長,然後再游回大海產卵。科學界早就知道生長在歐美淡水水域的鰻魚來自百慕達附近的馬尾藻海域,但沒人曉得日本鰻魚從哪裡來。
東京大學海洋研究所教授塚本勝巳領導的研究小組去年六月搭乘海洋研究船「白鳳丸」航行到馬里亞納群島西方靠近關島的海域。他們在海面下五百公尺深處撒下大型浮游生物網,並於「駿河海山」西方七十到一百公里處捕獲四百隻孵化約二至五天的鰻魚幼苗。經DNA鑑定,這些鰻苗確定是日本鰻。
研究人員按海流速度和孵化時間推算,認為位於北緯一四度、東經一四二度海底的「駿河海山」應該就是鰻魚產卵地。產下的鰻魚卵孵化後,隨北赤道海流漂向西方,而後再隨黑潮北上,經三個月到半年,游到台灣、中國大陸和日本海岸,鰻魚幼苗在這段期間會長大成為幼鰻,牠們接著便溯游至河流及湖泊中成長。
塚本將這項研究成果發表於廿三日出刊的最新一期英國《自然》周刊。他表示,確認鰻魚的產卵地之後,才能釐清鰻魚的迴游途徑,這項研究有助於保護日益稀少的日本鰻魚。
Wednesday, February 22, 2006
掉書袋(三)
譏笑人喜歡引經據典,咬文嚼字的毛病。
十國春秋˙南唐書˙卷三十二˙彭利用傳:對家人稚子,必據書史,斷章破句,以代常譚,俗謂之掉書袋。
解釋之
鰻魚養殖之成本,鰻苗支出佔五成左右。另一方面,由於直到目前鰻魚人工繁殖仍未成功,鰻苗來源完全依靠天然集捕。是故鰻苗供應的不確定及導致的價格波動,成為鰻魚工業發展的限制因子。人工飼養之鰻魚或野生鰻魚尚無在人為環境下自然性成熟之紀錄。而目前鰻魚人工受精卵之取得,皆由長期注射異種生物激素而達到。根據學者報告指出,人工激素 (異源性腦下垂體萃取液) 注射, 對性腺發育的效果,銀鰻的反應比黃鰻的好 (以卵巢濾泡細胞的發育情形或以卵的受精率及孵化率結果) (圖一)。顯示鰻魚其成長階段與激素的果效有特定之關係,代表銀化 (silvering) 為鰻魚青春期啟動 (onset of puberty) 的必要步驟。
銀化可視為鰻魚的第二次變態。 銀鰻和黃鰻的不同,在型態構造上包括: 背部含較多的黑色素,腹部有嘌呤 (purine) 的沉積,使腹部呈現銀白色;表皮變厚;側線變得更明顯;胸鰭變大;吻部變短;眼徑變得更大,網膜錐狀色素細胞漸被桿狀色素細胞取代,吸光波長改變;魚鰾變大內壁變厚;GSI 增加;消化道體重比 (digestive tracts- somatic index, DSI) 變小;也包括:體脂肪含量增加;間腎組織及甲狀腺活動活絡; 鰓部鹽類細胞 (chloride cells) 增加,鈉-鉀ATP 脢 (Na+-K+ ATPase) 的活性增加。在生理變化方面,生殖相關激素 (gonadotropin, GtH; gonadotropin-releasing hormone, GnRH; androgens; estrodial, E2) 及成長相關激素 (growth hormone, GH; thyroxine, T3/T4) 也有明顯的消長;肝臟中的脂質含量與粒腺體 (mitochondria) 數目昇高。值得提到的一點,如果黃鰻和銀鰻分別注射腦下垂體研磨萃取液,再調查血液中性類固醇的消長變化,發現銀鰻血液中雄性素 (T) 與雌性素 (E2) 皆昇高,而黃鰻只有雌性素 (E2) 昇高。事實上,台大漁科所羅教授與水試所鹿港分所證明,在日本鰻的人工催熟過程中,腦下垂體研磨萃取液中添加雄性素 (T) 已被證明有益於雌鰻配子發育的一致性。故其證明: 一、其所用之種鰻為黃鰻;二、前面所提的對性腺發育的效果,銀鰻的反應比黃鰻的好,雄性素 (T) 扮演某些重要功能。
雖然鰻魚的銀化對其降海生殖是必要的過程,但是到目前為止銀化如何調控,仍所知有限。事實上,已經證明可體松 (cortisol) 添加雄性素 (testosterone) 可刺激鰻魚銀化的某些特徵,如使黃鰻體表銀化,眼徑變大,消化道體重比變小,肝重體重比變大,最重要的是腦下垂體中的GtH-II 含量顯著提高 (圖二)。這一點是單用可體松或雄性素所無法達成的效果,事實上,此點有其生理的意義,因鰻魚為適應海水環境,鰓部鹽類細胞必須增加,鈉-鉀ATP 脢的活性也要加強,而可體松以經被證明有這些功效。最近荷蘭的調查報告指出,鰻魚在降海過程中血液中的可體松濃度有昇空的現象,更確定了其在鰻魚銀化的功能。附帶一點:一方面可體松是壓力激素 (stress hormone),對生殖有抑制作用;但另一方面,卻又對生殖激素 (GtH) 的生成有刺激作用 (看上去似乎矛盾,事實不然,它有它的演化和生理意義,以後再專文介紹之)。但要注意的是,可體松添加雄性素並無法刺激所有的銀化特徵,其它因子有待進一步的研究。
銀鰻被認為是處於準備進入 '青春期' (puberty) 的生活階段,外觀型態及行為的改變是經由內部的成長整合神經及內分泌變化而來。由這些身體變化特徵來看,銀化 (silvering) 可能是為了適應海洋生殖洄游做準備,因此也可將鰻魚的銀化視為準備青春期的開始 (preparation for the beginning of puberty),而銀鰻為青春期的開始 (the beginning of puberty)。但是如果被拘限在淡水環境中,銀鰻青春期啟動 (the onset of puberty) 卻是被抑制著的另一方面,以上所有的觀察結果及文獻几乎皆由歐洲鰻 (Anguilla anguilla) 的研究而來,亞洲特有之日本鰻 (Anguilla japonica) 相關之資料几乎付之闕如。
若鰻魚的銀化可視為準備青春期的開始,從生殖內分泌的觀點來看,最重要的是腦下垂體中的促性腺激素 (GtH) 的生成和釋放。因爲在所有的脊椎動物中,促性腺激素對性腺的發育和配子的生成扮演重要的角色,直接和生殖有關。同時青春期的發動與個體的年齡及成長的體型有關,因爲體型和體中堆積的能量有關,而母體若有足夠的能量累積,則可以確保母體生殖的成功和幼體發育的完整。 若銀化視為鰻魚的第二次變態,我想脊椎動物中,大家最熟悉的變態例子是蝌蚪變幼蛙與鮭魚的銀化 (smoltification)。在前一篇文章曾提到,和生長及新陳代謝有關的激素,可能在傳統的生殖激素外,對生殖及青春期的啟動扮演重要的功能,其中因素林 (insulin, 胰島素) 已經被證明和蝌蚪變態與鮭魚的銀化 (smoltification) 有關。
對鰻魚人工催熟而言,腦下垂體激素人工注射處理對卵巢發育的效果,銀鰻的結果比黃鰻顯著。顯示黃鰻體中欠缺某些與生殖密切連接之內在因子,這些因子可能是以化學訊息的方式存在 (如:神經傳遞物質, 神經內分泌或內分泌激素與某些堆積的能量物質)。故理論上可以利用人工方法,加速養殖鰻魚之銀化,達到穩定供應銀鰻,供應人工繁殖試驗之需求。至於養殖鰻要幾年才能答到銀鰻的標準呢?這個問題有賴大家一起的努力。
主要參考文獻:
Fontaine Y-A. (1994) L'argenture de l'anguille: metamorphose, anticipation, adaptation. Bull. Fr. Peche Piscic., 335: 171-185.
Huang Y-S., Schmitz M., Le Belle N., Chang C-F., Querat B. and Dufour S. (1997) Androgens stimulates gonadotropin-II subunit in eel pituitary cells in vitro. Mol. Cell. Endocrinol. 131:157-166.
Huang Y-S. (1998) Role des steroides sexuels et des hormones metaboliques dans le controle direct hypophysaire de l’hormone gonadotrope (GtH-II) chez l’anguille europeenne, Anguilla anguilla. Universite Paris VI, Departement de Physiologie de la Reproduction, 173p.
Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Schmitz M. and Dufour S. (1999) Cortisol selectively stimulates pituitary gonadotropin (GtH-II) subunit, in a primitive teleost, Anguilla anguilla. Endocrinology 140:1228-1235.
Ijiri S., KayabaT., Takeda N., Tachiki H., Adachi S. and Yamauchi K. (1998) Pretreatment reproductive stage and oocyte development induced by salmon pituitary homogenate in the Japanese eel Anguilla japonica. Fish.Sci., 62(1): 28-34.
十國春秋˙南唐書˙卷三十二˙彭利用傳:對家人稚子,必據書史,斷章破句,以代常譚,俗謂之掉書袋。
解釋之
鰻魚養殖之成本,鰻苗支出佔五成左右。另一方面,由於直到目前鰻魚人工繁殖仍未成功,鰻苗來源完全依靠天然集捕。是故鰻苗供應的不確定及導致的價格波動,成為鰻魚工業發展的限制因子。人工飼養之鰻魚或野生鰻魚尚無在人為環境下自然性成熟之紀錄。而目前鰻魚人工受精卵之取得,皆由長期注射異種生物激素而達到。根據學者報告指出,人工激素 (異源性腦下垂體萃取液) 注射, 對性腺發育的效果,銀鰻的反應比黃鰻的好 (以卵巢濾泡細胞的發育情形或以卵的受精率及孵化率結果) (圖一)。顯示鰻魚其成長階段與激素的果效有特定之關係,代表銀化 (silvering) 為鰻魚青春期啟動 (onset of puberty) 的必要步驟。
銀化可視為鰻魚的第二次變態。 銀鰻和黃鰻的不同,在型態構造上包括: 背部含較多的黑色素,腹部有嘌呤 (purine) 的沉積,使腹部呈現銀白色;表皮變厚;側線變得更明顯;胸鰭變大;吻部變短;眼徑變得更大,網膜錐狀色素細胞漸被桿狀色素細胞取代,吸光波長改變;魚鰾變大內壁變厚;GSI 增加;消化道體重比 (digestive tracts- somatic index, DSI) 變小;也包括:體脂肪含量增加;間腎組織及甲狀腺活動活絡; 鰓部鹽類細胞 (chloride cells) 增加,鈉-鉀ATP 脢 (Na+-K+ ATPase) 的活性增加。在生理變化方面,生殖相關激素 (gonadotropin, GtH; gonadotropin-releasing hormone, GnRH; androgens; estrodial, E2) 及成長相關激素 (growth hormone, GH; thyroxine, T3/T4) 也有明顯的消長;肝臟中的脂質含量與粒腺體 (mitochondria) 數目昇高。值得提到的一點,如果黃鰻和銀鰻分別注射腦下垂體研磨萃取液,再調查血液中性類固醇的消長變化,發現銀鰻血液中雄性素 (T) 與雌性素 (E2) 皆昇高,而黃鰻只有雌性素 (E2) 昇高。事實上,台大漁科所羅教授與水試所鹿港分所證明,在日本鰻的人工催熟過程中,腦下垂體研磨萃取液中添加雄性素 (T) 已被證明有益於雌鰻配子發育的一致性。故其證明: 一、其所用之種鰻為黃鰻;二、前面所提的對性腺發育的效果,銀鰻的反應比黃鰻的好,雄性素 (T) 扮演某些重要功能。
雖然鰻魚的銀化對其降海生殖是必要的過程,但是到目前為止銀化如何調控,仍所知有限。事實上,已經證明可體松 (cortisol) 添加雄性素 (testosterone) 可刺激鰻魚銀化的某些特徵,如使黃鰻體表銀化,眼徑變大,消化道體重比變小,肝重體重比變大,最重要的是腦下垂體中的GtH-II 含量顯著提高 (圖二)。這一點是單用可體松或雄性素所無法達成的效果,事實上,此點有其生理的意義,因鰻魚為適應海水環境,鰓部鹽類細胞必須增加,鈉-鉀ATP 脢的活性也要加強,而可體松以經被證明有這些功效。最近荷蘭的調查報告指出,鰻魚在降海過程中血液中的可體松濃度有昇空的現象,更確定了其在鰻魚銀化的功能。附帶一點:一方面可體松是壓力激素 (stress hormone),對生殖有抑制作用;但另一方面,卻又對生殖激素 (GtH) 的生成有刺激作用 (看上去似乎矛盾,事實不然,它有它的演化和生理意義,以後再專文介紹之)。但要注意的是,可體松添加雄性素並無法刺激所有的銀化特徵,其它因子有待進一步的研究。
銀鰻被認為是處於準備進入 '青春期' (puberty) 的生活階段,外觀型態及行為的改變是經由內部的成長整合神經及內分泌變化而來。由這些身體變化特徵來看,銀化 (silvering) 可能是為了適應海洋生殖洄游做準備,因此也可將鰻魚的銀化視為準備青春期的開始 (preparation for the beginning of puberty),而銀鰻為青春期的開始 (the beginning of puberty)。但是如果被拘限在淡水環境中,銀鰻青春期啟動 (the onset of puberty) 卻是被抑制著的另一方面,以上所有的觀察結果及文獻几乎皆由歐洲鰻 (Anguilla anguilla) 的研究而來,亞洲特有之日本鰻 (Anguilla japonica) 相關之資料几乎付之闕如。
若鰻魚的銀化可視為準備青春期的開始,從生殖內分泌的觀點來看,最重要的是腦下垂體中的促性腺激素 (GtH) 的生成和釋放。因爲在所有的脊椎動物中,促性腺激素對性腺的發育和配子的生成扮演重要的角色,直接和生殖有關。同時青春期的發動與個體的年齡及成長的體型有關,因爲體型和體中堆積的能量有關,而母體若有足夠的能量累積,則可以確保母體生殖的成功和幼體發育的完整。 若銀化視為鰻魚的第二次變態,我想脊椎動物中,大家最熟悉的變態例子是蝌蚪變幼蛙與鮭魚的銀化 (smoltification)。在前一篇文章曾提到,和生長及新陳代謝有關的激素,可能在傳統的生殖激素外,對生殖及青春期的啟動扮演重要的功能,其中因素林 (insulin, 胰島素) 已經被證明和蝌蚪變態與鮭魚的銀化 (smoltification) 有關。
對鰻魚人工催熟而言,腦下垂體激素人工注射處理對卵巢發育的效果,銀鰻的結果比黃鰻顯著。顯示黃鰻體中欠缺某些與生殖密切連接之內在因子,這些因子可能是以化學訊息的方式存在 (如:神經傳遞物質, 神經內分泌或內分泌激素與某些堆積的能量物質)。故理論上可以利用人工方法,加速養殖鰻魚之銀化,達到穩定供應銀鰻,供應人工繁殖試驗之需求。至於養殖鰻要幾年才能答到銀鰻的標準呢?這個問題有賴大家一起的努力。
主要參考文獻:
Fontaine Y-A. (1994) L'argenture de l'anguille: metamorphose, anticipation, adaptation. Bull. Fr. Peche Piscic., 335: 171-185.
Huang Y-S., Schmitz M., Le Belle N., Chang C-F., Querat B. and Dufour S. (1997) Androgens stimulates gonadotropin-II subunit in eel pituitary cells in vitro. Mol. Cell. Endocrinol. 131:157-166.
Huang Y-S. (1998) Role des steroides sexuels et des hormones metaboliques dans le controle direct hypophysaire de l’hormone gonadotrope (GtH-II) chez l’anguille europeenne, Anguilla anguilla. Universite Paris VI, Departement de Physiologie de la Reproduction, 173p.
Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Schmitz M. and Dufour S. (1999) Cortisol selectively stimulates pituitary gonadotropin (GtH-II) subunit, in a primitive teleost, Anguilla anguilla. Endocrinology 140:1228-1235.
Ijiri S., KayabaT., Takeda N., Tachiki H., Adachi S. and Yamauchi K. (1998) Pretreatment reproductive stage and oocyte development induced by salmon pituitary homogenate in the Japanese eel Anguilla japonica. Fish.Sci., 62(1): 28-34.
Tuesday, February 21, 2006
掉書袋(二)...
鰻魚爲亞洲之重要之經濟養殖魚種之一。但鰻魚的人工繁殖最大的問題之一是:其腦下垂體中的促性腺激素 (gonadotropin ,GtH) 含量相當的低,故目前鰻魚人工繁殖,其受精卵之取得,皆由長期注射異種生物腦下垂體研磨萃取液 (內含促性腺激素) 而來。早期魚類人工繁殖的實施也是利用注射腦下垂體研磨萃取液來達成,雖然兩者皆借用注射腦下垂體研磨萃取液,但其意義不同。對其他魚種而言,因其已經到達性熟,所欠缺的是引發堆積在腦下垂體中促性腺刺激素(GtH) 的高峰釋放 (discharge),進而引發配子的最後成熟 (final maturation) 與排卵 (ovulation) 及產卵 (spawning) 或排精 (spermation),因其配子以經發育到几乎完熟階段,故操作上可能只須施打一針。但對鰻魚而言,注射腦下垂體研磨萃取液,是要借著外來的激素,讓配子從原母細胞發育到完熟配子。故操作上需要施打數十針,時間也必須拉長。從上面所舉的例子,可看到促性腺激素 (gonadotropin ,GtH) 在魚類中至少有兩項功能:(一) 刺激配子發育,(二) 引發配子的最後成熟與產卵。但什麼是促性腺激素呢?在魚類有什麼特點呢?本文即在介紹有關魚類生殖內分泌的發展,著重於鰻魚腦下垂體中性腺刺激素的生理生化特性。
廣義而言:凡是可以刺激性腺活動的激素皆可稱為性腺促進激素 (gonadotropins, GtH)。這包含胎盤絨毛膜激素 (placenta-choronicgonadotropin, CG)、成長激素 (growth hormone, GH)、泌乳激素 (prolactin, PRL)、胰島素 (insulin) 類胰島素成長因子 (insulin-like-growth factors, IGF)…等等。但一般所指的 '性腺促進激素 (GtH)',是指由腦下垂體 (pituitary) 性腺促進激素細胞 (gonadotropes) 所產生的醣蛋白激素 (glycoprotein hormones)。
性腺促進激素 (GtH) 的物理化學特性,在哺乳動物中研究的比較清楚。在哺乳動物中有兩種性腺促進激素:包括濾泡刺激素 (follicle-stimulating hormone, FSH) 和黃體激素 (luteinizing hormone, LH),均為醣蛋白激素。在內分泌學上將甲狀腺刺激素 (thyroidotropin, TSH) 與 FSH、LH 歸為一大類,因為他們皆由兩個次單元體所構成:即 alpha 次單元體和 beta 次單元體,而FSH、LH、TSH 含有完全相同的alpha 次單元體,是 beta 次單元體的賦予各個激素不同的生理特性。但必須要兩個次單元體構成在一起才能具有生理活性。alpha 次單元體來自同一基因 (gene),而beta 次單元體由不同基因而來。
哺乳動物,在雌性中FSH 主要扮演刺激卵巢濾泡生長及芳香脢 (aromatase, 將睪固酮, testosterone, 轉變為雌性素, estrogens, 的酵素) 的活性;LH 則主要扮演刺激卵巢固醇類激素的生成 (主要是助孕素, progesterone,和雄性素,androgens) 刺激排卵與維持卵巢濾泡排卵後黃體 (corpus luteum) 的發育。而FSH 的受體 (receptors) 主要位於濾泡上的顆粒細胞 (granulosa cells) 層;LH 的受體主要位於卵巢的結構組織細胞 (stroma cells) 及濾泡上的鞘細胞 (theca cells) 和顆粒細胞 (granulosa cells) 層。雄性中,FSH 刺激施氏細胞 (Sertoli cells) 促進精子的生成;LH 刺激萊氏細胞 (Leydig cells) 促進雄性素的生成。故對哺乳動物的生殖而言,FSH 和 LH 的作用是相輔相乘的,雖各有特性但不能分開。
在魚類中,只存在著單一型式的促性腺激素 (GtH),亦或像哺乳動物一樣具有兩種不同促性腺激素 (即濾泡刺激素 FSH 和黃體激素LH),一直是個困惑的問題。最先在1971年,法國學者從鯉魚 (common carp) 的腦下垂體中分離出來一種物質,此物質具有雙重促性腺激素 (FSH 和LH) 的功能。不論在活體或離體實驗條件下均可刺激不同魚種如,金魚、鯰魚及鮭魚的性纇固醇激素合成和配子的最後成熟;同時證明它是一種醣蛋白,並由兩個次單元體所構成,這兩個次單元體被命名為次單元體-I (subunit-I) 及 單元體-II (subunit-II)。次單元體之間的結合和哺乳動物的情形一樣,並非靠著分子鍵結合。這些證據使他們推論在硬骨魚中可能只有一種促性腺激素。另外從氨基酸構成順序比較得知,單元體-I 的一級結構與哺乳動物促性腺激素的alpha次單元體的一級結構較相近,次單元體-II 的一級結構則和 LH 的 beta 次單元體的一級結構比較相近。
由哺乳動物而來的概念,一直有學者想證明在魚類中也具有兩種不同的GtH。在鮭魚類中也有一種促性腺激素被分離出來,比較詭弔的是几乎在卵巢發育的最後階段才能在血漿中測到這種到這種促性腺激素。因此,有人推測一定有另一種促性腺激素負責卵巢發育的前半部、但一直沒證據。直到 1988 年,兩種不同的促性腺激素 (GtH-I 和GtH-II) 才在鮭魚腦下垂體中被日本人分離及證明,鮭魚GtH-II 和先前從鯉魚所得到的 GtH 分子類似。到目前為止,兩種不同的GtH 已經在數十種魚類 (包括鯉魚、鰻魚) 中被確定,但特別的是,非洲鯰魚 (African catfish ) 到目前一直還沒找到。或許有人會問為什麼從1971年之前到 1988年才確定了魚類中也具有兩種不同的GtH,我想最大的緣因是因為分析技術的進步及分子生物學技術的廣汎應用,而GtH-I 生理功能的模糊性也是其難以肯定的原因之一。
由氨基酸一級結構比較顯示,不同魚類之間,其GtH-II 的beta 次單元體有較高的相似性;相反的,不同魚類之間其GtH-I 的beta 次單元體彼此有較大的差異。如果和高等脊椎動物的 GtH (LH, FSH) 相比較則發現,硬骨魚類GtH-II 的beta 次單元體和 LH 的beta 次單元體相近,而GtH-I 的beta 次單元體和 FSH 的beta 次單元體有較相近的氨基酸一級結構 (圖一)。值得一提的是,胎盤絨毛膜激素 (placenta-choronicgonadotropin, CG),其結構和功能則與LH相近,故早期養殖上,利用腦下垂體研磨萃取液添加人類胎盤絨毛膜激素 (hCG),作為魚類催熟的用藥,其原理是以hCG 代替魚類的GtH II (hCG = LH = GtH-II)。魚類GtH 和哺乳類動物GtH最大的不同的是:魚類GtH-I 據有兩種不同型式的 alpha 次單原體(alpha 1 和alpha 2)。
自從兩種不同的GtH 在多數的硬骨魚中被確立之後,大家更進一步的想到去了解它們的生理功能及被調節的機制,到目前為止,只有在鮭魚類中被研究得比較仔細而且具有較明顯的結果。利用高特異性的放射線免疫分析法(radioimmunoassay, RIA) 分別去偵測GtH-I 和GtH-II 在血液中變化的情形,發現在生殖季節時在配子生成階段血液中GtH-I 的濃度比 GtH-II 高出許多;但在此之後GtH-I 的濃度逐漸下降而GtH-II 的濃度逐漸上升。而在最後成熟階段時血液中GtH-II 有一高峰出現。推測不同的GtH 具有不同的生理功能,GtH-I 可能和配子生成及發育有關,而GtH-II 和配子最後成熟有關。此點 GtH-I 與GtH-II和 FSH與 LH 具有對映的相似功能。
至於在其他硬骨魚類 GtH-I 和GtH-II 就沒有如此清晰的出現型式,一方面因 GtH-I 和GtH-II相比較,其量本來就不多無法發展出完整的RIA系統,故現有的研究大多以偵測腦下垂體中GtH-I beta 和GtH-II beta 個別傳信核醣核酸 (mRNA) 的量。以金魚及鯛魚為例,GtH-I beta和 GtH-II beta的量,在卵子發育過程中兩者一起出現,並無顯著的差別;推測GtH-I 和GtH-II 對卵子發育可能具有同等的生理功能。對於某些缺乏GtH-II 的魚種,例如非洲鯰魚,GtH-II beta則出現在整個生殖季節。
若從受體的觀點來看,在鮭魚中發現兩種GtH 受體 (GtH-R)。其中 GtH-一型受體 (GtH-R-I) 可能和GtH-I 和GtH-II 結合,而GtH-二型受體 (GtH-R-II) 只能和GtH-II 結合。所以由不具GtH-I 專一性的受體 (GtH-R-I與GtH-R-II) 及GtH-I 的結構與出現在不同的魚種有很大的差別,推測在演化的過程中GtH-I 比GtH-II 晚出現。
這一點推測可能和鰻魚有關,鰻魚算是一種相當原始的魚類,雖然鰻魚的GtH 早在1983 年之前就被確立, 與鮭魚 GtH (GtH-I 和GtH-II) 比對結果,鰻魚的GtH應歸於 GtH-II 。從那時以後,法國及日本學者也紛紛加入尋找鰻魚GtH-I的行列,因為前面提到不同的GtH 具有不同的生理功能,GtH-I 可能和配子生成及發育有關,而GtH-II 和配子最後成熟有關。鰻魚所缺乏的是缺少刺激配子生成及發育的激素 (如GtH-I)。事實上,用傳統的內分泌技術,鰻魚和非洲鯰魚一樣根本測不到GtH-I的存在,法國派的學者比較趨向鰻魚只有一種 GtH (GtH-II) 的看法,但並不否認可能有 GtH-I 的存在,因為從雄鰻來的觀察顯示只要一針的LH 或 hCG 就可以使雄鰻達到排精的階段,顯示一種GtH 就能發揮所有生理功能。一直到1998年,日本學者以分子生物的技術 (RT-PCR) 終於在日本鰻證明GtH-I的存在,但是比較詭異的是,日本鰻的GtH-I只在小鰻才找得到而且量極少,催熟後的鰻魚 (不管雌雄鰻) 就不見了 (須知鰻魚在催熟後卵巢也才會發育,同時GtH-II的量也會昇高),顯示 GtH-I 可能和配子的生成及發育無直接關係。有可能鰻魚的GtH內分泌模式,介於非洲鯰魚和金魚之間,故GtH-I 對鰻魚可能是演化上進化的新分枝,但反過來說,也可能是退化的遺跡,更大膽的假設是:淡水或養殖鰻魚性成熟之所以受阻礙,乃是其GtH-I 的生成受阻,而催熟可能是一種病態的刺激才使 GtH-I 的生成受到抑制。事實上,傳統的魚類生殖內分泌對 GtH-II 的生成與調節研究的比較透澈,結果顯示,和哺乳動物動一樣受性腺激素的正負迴饋控制,是很古典的生殖內分泌,而且雖然魚種不同但有相同的結果。而對 GtH-I 的研究則剛開始起步,現有的結果顯示魚類GtH-I 的生成調控,隨著魚種的不同而有相反的結果 (以後再專文討論之)。
如果鰻魚的 GtH-I 是脊椎動物GtH系統 (GtH-system) 演化上進化的新分枝 (或是退化的遺跡),那麼如何以一種GtH去發揮所有的功能呢?有兩種可能性:一是,受體調節型:受體的觀點來看,可能鰻魚GtH-II同時可以和類似GtH-I 與GtH-II的受體作用,發揮GtH-I 與 GtH-II 的功能。二是,激素產生調節型:因為在哺乳動物中FSH和LH的生成與分泌各受不同的調控,如FSH的生成受雄性素、助孕素與可體松的刺激,但這些固醇類激素對LH無顯著刺激效果,特別的是,兩種性腺所產生的蛋白質激素稱為 activin和 inhibin,對FSH有非常特異的刺激或抑制效果,也就是說activin和 inhibin對LH無任何效果。事實上,從最近的實驗結果顯示,鰻魚的GtH-II具有和FSH類似的被調節機制,也就是說受雄性素、助孕素與可體松的刺激,而且至少inhibin對鰻魚的GtH-II有抑制效果 (但要記得的是,鰻魚的GtH-II它的構造與功能和LH較相近) 推測鰻魚的GtH是一種過渡型,即FSH加LH,即FSH 的操縱區 (promoter) 加上 LH 的構造基因 (structure gene)。這一點其中所含的生理及生物演化意義,值得大家再深入探究,有可能和鰻魚的生殖之謎有關,歡迎大家一起集思廣益。
本系列文章下回將與大家報告所謂的黃鰻,銀鰻的差別,及其生理意義,與人工催熟對其的不同效果。
主要參考文獻:
Querat B. (1994) Molecular evolution of the glycoprotein hormones in vertebrates. In: Davey K.G., Peter R. E. and Tope S.S. (Eds) 'Persperctives in Comparative Endocrinology', NRC, Toronto, Ontario, Canada. pp27-35.
Huang Y-S., Schmitz M., Le Belle N., Chang C-F., Querat B. and Dufour S. (1997) Androgens stimulates gonadotropin-II subunit in eel pituitary cells in vitro. Mol. Cell. Endocrinol. 131:157-166.
Huang Y-S., Le Belle N., Schmitz M., Chang C-F., Vernier P., Querat B. and Dufour S. (1998) In vivo and in vitro effects of sex steroids on gonadotropin (GtH-II) synthesis in the female European eel. Trends Comp. Endocrinol. Neurobiol., Ann. New York Acad. Sci. 839:348-350.
Huang Y-S. (1998) Role des steroides sexuels et des hormones metaboliques dans le controle direct hypophysaire de l’hormone gonadotrope (GtH-II) chez l’anguille europeenne, Anguilla anguilla. Universite Paris VI, Departement de Physiologie de la Reproduction, 173p.
Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Schmitz M. and Dufour S. (1999) Cortisol selectively stimulates pituitary gonadotropin (GtH-II) subunit, in a primitive teleost, Anguilla anguilla. Endocrinology 140:1228-1235.
廣義而言:凡是可以刺激性腺活動的激素皆可稱為性腺促進激素 (gonadotropins, GtH)。這包含胎盤絨毛膜激素 (placenta-choronicgonadotropin, CG)、成長激素 (growth hormone, GH)、泌乳激素 (prolactin, PRL)、胰島素 (insulin) 類胰島素成長因子 (insulin-like-growth factors, IGF)…等等。但一般所指的 '性腺促進激素 (GtH)',是指由腦下垂體 (pituitary) 性腺促進激素細胞 (gonadotropes) 所產生的醣蛋白激素 (glycoprotein hormones)。
性腺促進激素 (GtH) 的物理化學特性,在哺乳動物中研究的比較清楚。在哺乳動物中有兩種性腺促進激素:包括濾泡刺激素 (follicle-stimulating hormone, FSH) 和黃體激素 (luteinizing hormone, LH),均為醣蛋白激素。在內分泌學上將甲狀腺刺激素 (thyroidotropin, TSH) 與 FSH、LH 歸為一大類,因為他們皆由兩個次單元體所構成:即 alpha 次單元體和 beta 次單元體,而FSH、LH、TSH 含有完全相同的alpha 次單元體,是 beta 次單元體的賦予各個激素不同的生理特性。但必須要兩個次單元體構成在一起才能具有生理活性。alpha 次單元體來自同一基因 (gene),而beta 次單元體由不同基因而來。
哺乳動物,在雌性中FSH 主要扮演刺激卵巢濾泡生長及芳香脢 (aromatase, 將睪固酮, testosterone, 轉變為雌性素, estrogens, 的酵素) 的活性;LH 則主要扮演刺激卵巢固醇類激素的生成 (主要是助孕素, progesterone,和雄性素,androgens) 刺激排卵與維持卵巢濾泡排卵後黃體 (corpus luteum) 的發育。而FSH 的受體 (receptors) 主要位於濾泡上的顆粒細胞 (granulosa cells) 層;LH 的受體主要位於卵巢的結構組織細胞 (stroma cells) 及濾泡上的鞘細胞 (theca cells) 和顆粒細胞 (granulosa cells) 層。雄性中,FSH 刺激施氏細胞 (Sertoli cells) 促進精子的生成;LH 刺激萊氏細胞 (Leydig cells) 促進雄性素的生成。故對哺乳動物的生殖而言,FSH 和 LH 的作用是相輔相乘的,雖各有特性但不能分開。
在魚類中,只存在著單一型式的促性腺激素 (GtH),亦或像哺乳動物一樣具有兩種不同促性腺激素 (即濾泡刺激素 FSH 和黃體激素LH),一直是個困惑的問題。最先在1971年,法國學者從鯉魚 (common carp) 的腦下垂體中分離出來一種物質,此物質具有雙重促性腺激素 (FSH 和LH) 的功能。不論在活體或離體實驗條件下均可刺激不同魚種如,金魚、鯰魚及鮭魚的性纇固醇激素合成和配子的最後成熟;同時證明它是一種醣蛋白,並由兩個次單元體所構成,這兩個次單元體被命名為次單元體-I (subunit-I) 及 單元體-II (subunit-II)。次單元體之間的結合和哺乳動物的情形一樣,並非靠著分子鍵結合。這些證據使他們推論在硬骨魚中可能只有一種促性腺激素。另外從氨基酸構成順序比較得知,單元體-I 的一級結構與哺乳動物促性腺激素的alpha次單元體的一級結構較相近,次單元體-II 的一級結構則和 LH 的 beta 次單元體的一級結構比較相近。
由哺乳動物而來的概念,一直有學者想證明在魚類中也具有兩種不同的GtH。在鮭魚類中也有一種促性腺激素被分離出來,比較詭弔的是几乎在卵巢發育的最後階段才能在血漿中測到這種到這種促性腺激素。因此,有人推測一定有另一種促性腺激素負責卵巢發育的前半部、但一直沒證據。直到 1988 年,兩種不同的促性腺激素 (GtH-I 和GtH-II) 才在鮭魚腦下垂體中被日本人分離及證明,鮭魚GtH-II 和先前從鯉魚所得到的 GtH 分子類似。到目前為止,兩種不同的GtH 已經在數十種魚類 (包括鯉魚、鰻魚) 中被確定,但特別的是,非洲鯰魚 (African catfish ) 到目前一直還沒找到。或許有人會問為什麼從1971年之前到 1988年才確定了魚類中也具有兩種不同的GtH,我想最大的緣因是因為分析技術的進步及分子生物學技術的廣汎應用,而GtH-I 生理功能的模糊性也是其難以肯定的原因之一。
由氨基酸一級結構比較顯示,不同魚類之間,其GtH-II 的beta 次單元體有較高的相似性;相反的,不同魚類之間其GtH-I 的beta 次單元體彼此有較大的差異。如果和高等脊椎動物的 GtH (LH, FSH) 相比較則發現,硬骨魚類GtH-II 的beta 次單元體和 LH 的beta 次單元體相近,而GtH-I 的beta 次單元體和 FSH 的beta 次單元體有較相近的氨基酸一級結構 (圖一)。值得一提的是,胎盤絨毛膜激素 (placenta-choronicgonadotropin, CG),其結構和功能則與LH相近,故早期養殖上,利用腦下垂體研磨萃取液添加人類胎盤絨毛膜激素 (hCG),作為魚類催熟的用藥,其原理是以hCG 代替魚類的GtH II (hCG = LH = GtH-II)。魚類GtH 和哺乳類動物GtH最大的不同的是:魚類GtH-I 據有兩種不同型式的 alpha 次單原體(alpha 1 和alpha 2)。
自從兩種不同的GtH 在多數的硬骨魚中被確立之後,大家更進一步的想到去了解它們的生理功能及被調節的機制,到目前為止,只有在鮭魚類中被研究得比較仔細而且具有較明顯的結果。利用高特異性的放射線免疫分析法(radioimmunoassay, RIA) 分別去偵測GtH-I 和GtH-II 在血液中變化的情形,發現在生殖季節時在配子生成階段血液中GtH-I 的濃度比 GtH-II 高出許多;但在此之後GtH-I 的濃度逐漸下降而GtH-II 的濃度逐漸上升。而在最後成熟階段時血液中GtH-II 有一高峰出現。推測不同的GtH 具有不同的生理功能,GtH-I 可能和配子生成及發育有關,而GtH-II 和配子最後成熟有關。此點 GtH-I 與GtH-II和 FSH與 LH 具有對映的相似功能。
至於在其他硬骨魚類 GtH-I 和GtH-II 就沒有如此清晰的出現型式,一方面因 GtH-I 和GtH-II相比較,其量本來就不多無法發展出完整的RIA系統,故現有的研究大多以偵測腦下垂體中GtH-I beta 和GtH-II beta 個別傳信核醣核酸 (mRNA) 的量。以金魚及鯛魚為例,GtH-I beta和 GtH-II beta的量,在卵子發育過程中兩者一起出現,並無顯著的差別;推測GtH-I 和GtH-II 對卵子發育可能具有同等的生理功能。對於某些缺乏GtH-II 的魚種,例如非洲鯰魚,GtH-II beta則出現在整個生殖季節。
若從受體的觀點來看,在鮭魚中發現兩種GtH 受體 (GtH-R)。其中 GtH-一型受體 (GtH-R-I) 可能和GtH-I 和GtH-II 結合,而GtH-二型受體 (GtH-R-II) 只能和GtH-II 結合。所以由不具GtH-I 專一性的受體 (GtH-R-I與GtH-R-II) 及GtH-I 的結構與出現在不同的魚種有很大的差別,推測在演化的過程中GtH-I 比GtH-II 晚出現。
這一點推測可能和鰻魚有關,鰻魚算是一種相當原始的魚類,雖然鰻魚的GtH 早在1983 年之前就被確立, 與鮭魚 GtH (GtH-I 和GtH-II) 比對結果,鰻魚的GtH應歸於 GtH-II 。從那時以後,法國及日本學者也紛紛加入尋找鰻魚GtH-I的行列,因為前面提到不同的GtH 具有不同的生理功能,GtH-I 可能和配子生成及發育有關,而GtH-II 和配子最後成熟有關。鰻魚所缺乏的是缺少刺激配子生成及發育的激素 (如GtH-I)。事實上,用傳統的內分泌技術,鰻魚和非洲鯰魚一樣根本測不到GtH-I的存在,法國派的學者比較趨向鰻魚只有一種 GtH (GtH-II) 的看法,但並不否認可能有 GtH-I 的存在,因為從雄鰻來的觀察顯示只要一針的LH 或 hCG 就可以使雄鰻達到排精的階段,顯示一種GtH 就能發揮所有生理功能。一直到1998年,日本學者以分子生物的技術 (RT-PCR) 終於在日本鰻證明GtH-I的存在,但是比較詭異的是,日本鰻的GtH-I只在小鰻才找得到而且量極少,催熟後的鰻魚 (不管雌雄鰻) 就不見了 (須知鰻魚在催熟後卵巢也才會發育,同時GtH-II的量也會昇高),顯示 GtH-I 可能和配子的生成及發育無直接關係。有可能鰻魚的GtH內分泌模式,介於非洲鯰魚和金魚之間,故GtH-I 對鰻魚可能是演化上進化的新分枝,但反過來說,也可能是退化的遺跡,更大膽的假設是:淡水或養殖鰻魚性成熟之所以受阻礙,乃是其GtH-I 的生成受阻,而催熟可能是一種病態的刺激才使 GtH-I 的生成受到抑制。事實上,傳統的魚類生殖內分泌對 GtH-II 的生成與調節研究的比較透澈,結果顯示,和哺乳動物動一樣受性腺激素的正負迴饋控制,是很古典的生殖內分泌,而且雖然魚種不同但有相同的結果。而對 GtH-I 的研究則剛開始起步,現有的結果顯示魚類GtH-I 的生成調控,隨著魚種的不同而有相反的結果 (以後再專文討論之)。
如果鰻魚的 GtH-I 是脊椎動物GtH系統 (GtH-system) 演化上進化的新分枝 (或是退化的遺跡),那麼如何以一種GtH去發揮所有的功能呢?有兩種可能性:一是,受體調節型:受體的觀點來看,可能鰻魚GtH-II同時可以和類似GtH-I 與GtH-II的受體作用,發揮GtH-I 與 GtH-II 的功能。二是,激素產生調節型:因為在哺乳動物中FSH和LH的生成與分泌各受不同的調控,如FSH的生成受雄性素、助孕素與可體松的刺激,但這些固醇類激素對LH無顯著刺激效果,特別的是,兩種性腺所產生的蛋白質激素稱為 activin和 inhibin,對FSH有非常特異的刺激或抑制效果,也就是說activin和 inhibin對LH無任何效果。事實上,從最近的實驗結果顯示,鰻魚的GtH-II具有和FSH類似的被調節機制,也就是說受雄性素、助孕素與可體松的刺激,而且至少inhibin對鰻魚的GtH-II有抑制效果 (但要記得的是,鰻魚的GtH-II它的構造與功能和LH較相近) 推測鰻魚的GtH是一種過渡型,即FSH加LH,即FSH 的操縱區 (promoter) 加上 LH 的構造基因 (structure gene)。這一點其中所含的生理及生物演化意義,值得大家再深入探究,有可能和鰻魚的生殖之謎有關,歡迎大家一起集思廣益。
本系列文章下回將與大家報告所謂的黃鰻,銀鰻的差別,及其生理意義,與人工催熟對其的不同效果。
主要參考文獻:
Querat B. (1994) Molecular evolution of the glycoprotein hormones in vertebrates. In: Davey K.G., Peter R. E. and Tope S.S. (Eds) 'Persperctives in Comparative Endocrinology', NRC, Toronto, Ontario, Canada. pp27-35.
Huang Y-S., Schmitz M., Le Belle N., Chang C-F., Querat B. and Dufour S. (1997) Androgens stimulates gonadotropin-II subunit in eel pituitary cells in vitro. Mol. Cell. Endocrinol. 131:157-166.
Huang Y-S., Le Belle N., Schmitz M., Chang C-F., Vernier P., Querat B. and Dufour S. (1998) In vivo and in vitro effects of sex steroids on gonadotropin (GtH-II) synthesis in the female European eel. Trends Comp. Endocrinol. Neurobiol., Ann. New York Acad. Sci. 839:348-350.
Huang Y-S. (1998) Role des steroides sexuels et des hormones metaboliques dans le controle direct hypophysaire de l’hormone gonadotrope (GtH-II) chez l’anguille europeenne, Anguilla anguilla. Universite Paris VI, Departement de Physiologie de la Reproduction, 173p.
Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Schmitz M. and Dufour S. (1999) Cortisol selectively stimulates pituitary gonadotropin (GtH-II) subunit, in a primitive teleost, Anguilla anguilla. Endocrinology 140:1228-1235.
Monday, February 20, 2006
掉書袋...
英科學家稱尼斯湖水怪可能是100多歲的巨鰻
據倫敦9月23日(北京時間24日)消息,英國神秘動物學家理查德-弗裡曼在《太陽報》上發表了一份研究成果,認為著名的尼斯湖水怪可能是有著100多歲高齡、喪失了繁殖能力的巨鰻。
弗裡曼表示,這種鰻魚的身長在8-9米左右,獨特的自然條件十它得以長壽。他認為,實際上尼斯湖裡不只有一條這樣的鰻魚,應該存在著多條巨鰻。他還展示了在尼斯湖拍攝的一些照片作為證據。
弗裡曼說:『鰻魚一般在長到10歲左右就游到佛羅裡達沿岸海域繁衍,然後死亡。但某個因素對某種鰻魚產生了影響,致使其喪失繁衍能力,於是它們便不再前往大西洋繁衍後代,留在原地繼續生長,逐漸達到令人難以置信的龐大體積。』
他還指出,在尼斯湖裡存在著一種現象,導致鰻魚能夠以超出常理的速度生長,存活時間也大大拉長。
他認為,與關於尼斯湖水怪的其他說法相比,這是唯一有科學依據的解釋。
最新理論 尼斯湖水怪是幾條無法生育的〔太監〕鰻魚
由於鰻魚特殊的生活史, 如長達十數年的淡水生長期, 由鰻線, 幼鰻, 小鰻, 長成黃鰻 (yellow eel), 黃鰻再長成銀鰻 (silver eel), 在天然環境中, 所謂的黃鰻, 銀鰻是由其腹部的顔色而區分. 銀鰻仍處於性發育前期 (pre-puberty), 但如果不降海則永遠無法達到性成熟. 銀鰻在降海之後, 在海洋洄游數千公里到達深達百米以上的生殖産卵場. 以目前的科技水準, 仍對精確産卵場之判定有困難. 日本學者曾預測鰻魚在水深75-100米左右的水深孵化. 由人工繁殖之經驗推定, 鰻魚之受精卵應爲浮性卵, 卵徑約在 1mm左右, 可漂浮在 34 ppt 的海水中. 鰻魚孵化出來之後, 體型漸漸變成頭小身體透明扁平之柳葉狀, 故又稱柳葉鰻 (leptocephalus), 此體型構造爲外洋性浮游動植物之型態特徵之一. 此時柳葉鰻無自由行動之能力, 隨洋流漂浮至大陸邊緣, 依種類的不同, 此段時間預估約須一年以上, 在這段時間, 仔鰻之型態由柳葉狀變成長細型, 但仍爲透明故又稱玻璃鰻 (glass eel). 玻璃鰻已有自由行動之能力並對特定物質有驅近現象. 玻璃鰻在河口進入淡水後, 體表色素出現並長成爲小鰻. 由上述之鰻魚生活簡史, 可以知道鰻魚人工繁殖之困難點在: 之一, 影響鰻魚性成熟之內在 (例如: 內分泌, 體成份變化) 與環境 (例如: 鹽度, 溫度, 光照, 壓力…等等) 因子不明; 人工飼養之鰻魚或野生種鰻尚無在人爲環境下自然性成熟之紀錄. 之二, 仔鰻之孵化, 成長條件及初期餌料並不清楚, 若以一般海水魚類初期餌料, 如ss型海水輪蟲或戶外養成池池水, 最長僅可使幼苗存活一個月. 之三, 而目前鰻魚人工繁殖, 其受精卵之取得, 皆由長期注射異種生物激素而來, 使鰻魚處在不正常的生理狀況下; 報告指出, 既使在虹(魚尊)或鯉魚 使用人工催熟産生之幼苗, 比自然産卵受精孵化的幼苗, 具較高之死亡率和畸形率. 可見欲達成鰻魚人工繁殖之成功, 可能需由解決此三方面之問題齊下. 鰻魚的生殖生理的問題可能和其他魚類不同, 對水産養殖而言, 因爲其他魚種已經達性熟, 是欠缺引發堆積在腦下垂體中促性腺刺激素 (gonadotropins, GtHs) 的釋放 (discharge), 而引發配子的最後成熟. 所以所在早期水産養殖, 所謂的促性腺刺激素釋放激素的長效合成物 (gonadotropin-releasing hormone analogues, GnRH-a) 與多巴胺拮抗物 (anti-dopamine reagents) 有其應用上的價值. 而鰻魚的問題之一則是: 腦下垂體中的促性腺激素含量相當的低, 是故性腺刺激素釋放激素的長效合成物 (gonadotropin-releasing hormone analogues, GnRH-a) 或多巴胺拮抗物 (anti-dopamine reagents) 對鰻魚毫無效果的原因在此. 本文即在介紹有關鰻魚生殖內分泌的最新發展, 著重於腦下垂體中性腺刺激素生成的調控.
脊椎動物的生殖作用是由下視丘-腦下垂體-性腺所組成的神經內分泌軸線 (hypothalamus-pituitary-gonads neuroendocrine axis, HPG axis) 所控制. 腦部下視丘分泌的促性腺激素釋放激素 (gonadotropin-releasing hormone, GnRH) 控制腦下垂體中促性腺激素 (gonadotropins, 在高等脊椎動物是黃體激素, lutenizeing hormone, LH, 及濾泡刺激素, follicle-stimulating hormone, FSH; 在魚類則是促性腺刺激素一型, GtH-I, 及促性腺刺激素二型, GtH-II, 構造上 GtH-II 和 LH 較相近 而 GtH-I 則與 FSH 較相近, 其演化及生理意義以後再專文介紹之) 的生成和釋放; 而促性腺激素的作用則是刺激性腺的功能, 包括配子生成 (gametogenesis) 及類固醇激素的生成 (steroidogenesis). 在魚類, 促性腺激素的生成與釋放, 除了受促性腺激素釋激素 (GnRH) 的刺激外, 也同時由多巴胺 (dopamine) 所抑制. 而性腺所産生的各種激素 (如: 性類固醇激素), 能回饋控制腦部下視丘及腦下垂體, 進而影響達這條軸線的作用. 外部環境因子包括: 物理-化學, 營養及體內部狀況 (如: 生長及新陳代謝) 也能影響下視丘-腦下垂體-性腺神經內分泌軸的功能.
生物的發育並不是由稚體突然變成具生殖能力的成體, 而是需要 '青春期' 的過渡及準備. 所謂的 '青春期' (puberty) 是指生物稚體變成具有生殖能力成體的一段過程; 在這段過程中, 生物體會有某些改變, 包括: 外部型態, 生理狀況, 行爲…等等. 青春期的英文名辭 ‘puberty’ 是由 '體毛' (pubes) 這個拉丁字而來的, 指人類在青春期時某些部位的體毛(或體毛的生長). 後來泛指哺乳動物及其它脊椎動物, 包括: 鳥類及魚類的生殖發動準備期. 現在包括無脊椎動物, 都利用這個名辭.
在魚類, 青春期 (生殖發動準備期) 的認定, 在雄性, 爲精子 (spermatocyte) 的出現; 在雌性, 爲卵黃生成 (vitellogenesis) 的開始. 而青春期的完成則以第一次排卵或排精爲認定標準, 即個體具備生殖能力. 所以青春期是一個動態的演變現象, 在這一段時間內, 稚體完成具有生殖能力成體的所有準備. 從生殖內分泌的觀點來看, 最重要的是腦下垂體中的促性腺激素的生成和釋放, 因爲在所有的脊椎動物中, 促性腺激素對性腺的發育和配子的生成扮演重要的角色, 直接和生殖有關. 青春期的發動與個體的年齡及成長的體型有關, 因爲體型和體中堆積的能量有關, 而母體若有足夠的能量累積可, 則以確保母體生殖的成功和幼體發育的完整. 例如: 某些具生殖遷徙行爲的動物, 必須遷徙到適當環境, 才可能發生生殖行爲; 在卵生動物, 能量以卵黃由母體轉移到子代; 在哺乳動物, 母體能量經由胎盤及乳汁轉移到子代. 所以與生長及新陳代謝有關的激素, 可能在傳統的生殖激素外, 對生殖及青春期的啟動扮演重要的功能.
魚類和其他脊椎動物一樣, 已有有相當多的學術證據及現場經驗, 支持成長率和生殖啟動 (青春期) 的正相關性. 但到目前爲止, 卻只有很少的學術報告指明, 是何種化學訊息及生理機制連接體成長和青春期的啟動. 體成長主要受成表激素 (growth hormone, GH) 的調控, 但腦下垂體細胞培養實驗證明, 成長激素和促性腺激素的生成或釋放沒有直接的關係, 推測成長激素可能需要借由其他因子來連接體成長和生殖啟動. 最近的報告指出, 類胰島素成長因子 (insulin-like growth factors, IGFs) 在離體實驗條件下, 可促進鰻魚腦下垂體中促性腺激素的生成和釋放. 類胰島素成長因子的生成, 特別是類胰島素成長因子一型 (insulin-like growth factor-I, IGF-I), 受成長激素的控制, 主要的生成部在肝臟再釋放到血液循環中, 在其他組織中, 受成長激素的調控, 也有少量的區域性生成. 成長激素的作用需要類胰島素成長因子一型的協同作用, 才能發揮最大作用; 最明顯的例子, 魚類在饑餓狀態下, 爲調節能量平衡, 血液中有高量的成長激素, 可是此時成長卻是被抑制的, 同時類胰島素成長因子一型的血液含量也是很低. 而其他和生長及發育有關的激素, 例如甲狀腺素 (thyroxins) 已被證明對鰻魚腦下垂體中促性腺激素的生成無任何作用. 顯示類胰島素成長因子爲連結體成長和生殖啟動的化學訊息. 不止在鰻魚, 最近的報告也指出, 在鮭鱒魚類中, 類胰島素成長因子一型也可以刺激腦下垂體中促性腺激素的含量, 同時也能加強促性腺激素釋放激素對刺激腦下垂體性腺刺激素釋放的效能. 這一點在哺乳動中也有相同的結果. 以上的證據說明, 除了刺激體成長外, 類胰島素成長因子一型也可以直接刺激素腦下垂體中促性腺刺激素的生成和釋放, 更確定類胰島素成長因子一型是連接體成長和青春期 (生殖) 啟動的化學因數.
所謂良好的生殖體型包括: 體型大小及足夠的能量累積, 以應付在生殖過程中所需的能量. 和其他脊椎動物一樣, 胰島素 (insulin) 在魚類中, 對蛋白質和脂肪質的同化代謝作用扮演著極重要的功能. 在鮭魚的銀化 (smoltification) 和兩棲類的變態, 胰島素被證明具有觸發啟始的作用, 可能與其對能量轉換所具有的重要影響有關. 在鰻魚, 實驗證明胰島素的注射可顯著的提高鰻魚腦下垂體中促性腺激素的含量. 對鰻魚而言, 銀鰻 (成鰻) 比黃鰻 (小鰻) 體成份中含更高量的脂肪. 這種現象使學者提出脂質的堆積到某一個臨界量, 是推動鰻魚進入生殖發動期的重要因素, 這種概念在哺乳動物中早已經被提出. 在哺乳動物中, 目前能量堆積和生殖作用彼此交互影響的研究方面, 抑脂激素 (leptin) 是一個重點, 抑脂激素是由脂肪細胞所製造分泌, 它和食欲及攝食行爲有關. 目前有相當多的報告指出, 抑脂激素和促性腺激素釋放激素的作用有關, 而肥胖症和某些營養不良引起的不孕, 可用由抑脂素的處理而得到矯正. 雖然抑脂激素在魚類中的存在尚未被確定, 但是由分子生物技術確定在鰻魚中有此基因的存在. 同時, 銀鰻的體脂肪含量可高達 30% 以上, 抑脂激素的存在可能對鰻魚的生殖具有某些功能, 而離體腦下垂體細胞培養的實驗證明, 抑脂激素有促進鰻魚促性腺激素含量的功效, 同時也可刺激虹鱒促性腺激素的釋放. 由這些證據的支援了, 在魚類中體脂肪質的堆積和生殖發動的生理基礎, 可能是藉由胰島素和抑脂激素所傳遞的.
在青春期中, 特別是卵生動物, 身體所貯藏的能量必須轉移到性腺的發育 (包括配子細胞及性腺細胞本身的發育). 某些魚類, 例如鰻魚或鮭魚在生殖期時呈斷食的狀態而且需要進行長達數千公里的生殖回游, 這些能量均需在青春期中準備堆積好. 以鰻魚爲例, 在銀鰻階段時即開始生理性斷食, 而性成熟的雌魚其性腺體重比 (GSI) 可由原來的 2 % 成長到 50 % 以上. 而這現象是在鰻魚回游數千公里後發生的, 身體所貯藏的能量機乎完全轉移到性腺的發育. 可體素 (cortisol) 由其腎上腺皮質所製造分泌, 對生物的能量中間代謝有關包括屯積脂肪的運用, 及刺激醣質新生作用 (gluconeogenesis), 些作用在脊椎動物中是共通的. 在哺乳動物中, 可體素除了被證明和壓力的有關, 會抑制生殖作用之外, 同時也被證明和青春的啟動有關, 在魚類也有相同的報告, 抑制成魚生殖作用, 卻刺激稚魚, 包括虹鱒及鰻魚, 腦下垂體促性腺素的含量. 對鰻魚而言,活體和離體細胞腦下垂體細胞培養都證明了可體素具有刺激素腦下垂體促性腺素生成作用. 這些證據顯示可體素對生殖的作用, 除了在能量中間代謝層面外, 也可直接作用在腦下垂體刺激性腺刺激素的生成.
由上述的實驗中的結果說明, 魚類的腦下垂體中的促性腺激素細胞, 除了受傳統的下視丘-腦下垂體-性腺所組成的神經內分泌軸線所控制, 包括: 促性腺激素釋放激素, 多巴胺, 促性腺激素及性類固醇的調控之外, 某些所謂週邊訊息 (peripheral messengers) , 包括: 體成長, 新陳代謝情形, 組織貯能狀況, 可藉由如類成胰島素成長因子 (IGFs), 可體素 (cortisol), 胰島素 (insulin) 及抑脂激素 (leptin) 等而達到生殖調控的目的.
事實上大家早就了解種魚選擇的標準之一, 就是肥滿度, 包括體型及觸感所得的性腺發育情形. 而鰻魚, 特別是養殖鰻, 從生殖生理的觀點並無所謂的 '種鰻', 只有長得比較大的鰻.謹以此文拋磚引玉, 歡迎大家一起討論.
主要參考文獻:
- Dufour S., Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Vidal B., Marchelidon J., Querat B., Burzawa-Gerard E., Chang C-F., Schmitz M. (1999) Puberty in teleosts: new insights into the role of peripheral signals in the stimulation on pituitary gonadotropins. Proceedings of thr 6th International Symposium on the Reproductive Physiology of Fish, Bergen, Norway, July 4-9, 1999. (in press).
- Huang Y-S., Rousseau K., Le Belle N., Vidal B., Burzawa-Gerard E., Marchelidon J. and Dufour S. (1998) Insulin-like growth factor I (IGF-I) stimulates gonadotropin production from eel pituitary cells: a possible metabolic signal for induction of puberty. J. Endocrinol. 159:43-52.
- Huang Y-S. (1998) Role des steroides sexuels et des hormones metaboliques dans le controle direct hypophysaire de l’hormone gonadotrope (GtH-II) chez l’anguille europeenne, Anguilla anguilla. Universite Paris VI, Departement de Physiologie de la Reproduction, 173p.
- Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Schmitz M. and Dufour S. (1999) Cortisol selectively stimulates pituitary gonadotropin (GtH-II) subunit, in a primitive teleost, Anguilla anguilla. Endocrinology 140:1228-1235.
- Huang Y-S., Rousseau K., Le Belle N., Vidal B., Burzawa-Gerard E., Marchelidon J. and Dufour S. (1999) Opposite effects of insulin-like growth factors (IGFs) on gonadotropin (GtH-II) and growth hormone (GH) production by primary culture of European eel ( Anguilla anguilla ) pituitary cells. Aquaculture 177:73-83.
-Rousseau K., Huang Y-S., Le Belle N., Vidal B., Marchelidon J., Epelbaum J. and Dufour S. (1998) Long-term growth hormone release by serum-free primary culture of pituitary cells from European eel ( Anguilla anguilla ): inhibitory effects of somatostatin and insulin-like growth factors. Neuroendocrinology 67:301-309.
據倫敦9月23日(北京時間24日)消息,英國神秘動物學家理查德-弗裡曼在《太陽報》上發表了一份研究成果,認為著名的尼斯湖水怪可能是有著100多歲高齡、喪失了繁殖能力的巨鰻。
弗裡曼表示,這種鰻魚的身長在8-9米左右,獨特的自然條件十它得以長壽。他認為,實際上尼斯湖裡不只有一條這樣的鰻魚,應該存在著多條巨鰻。他還展示了在尼斯湖拍攝的一些照片作為證據。
弗裡曼說:『鰻魚一般在長到10歲左右就游到佛羅裡達沿岸海域繁衍,然後死亡。但某個因素對某種鰻魚產生了影響,致使其喪失繁衍能力,於是它們便不再前往大西洋繁衍後代,留在原地繼續生長,逐漸達到令人難以置信的龐大體積。』
他還指出,在尼斯湖裡存在著一種現象,導致鰻魚能夠以超出常理的速度生長,存活時間也大大拉長。
他認為,與關於尼斯湖水怪的其他說法相比,這是唯一有科學依據的解釋。
最新理論 尼斯湖水怪是幾條無法生育的〔太監〕鰻魚

由於鰻魚特殊的生活史, 如長達十數年的淡水生長期, 由鰻線, 幼鰻, 小鰻, 長成黃鰻 (yellow eel), 黃鰻再長成銀鰻 (silver eel), 在天然環境中, 所謂的黃鰻, 銀鰻是由其腹部的顔色而區分. 銀鰻仍處於性發育前期 (pre-puberty), 但如果不降海則永遠無法達到性成熟. 銀鰻在降海之後, 在海洋洄游數千公里到達深達百米以上的生殖産卵場. 以目前的科技水準, 仍對精確産卵場之判定有困難. 日本學者曾預測鰻魚在水深75-100米左右的水深孵化. 由人工繁殖之經驗推定, 鰻魚之受精卵應爲浮性卵, 卵徑約在 1mm左右, 可漂浮在 34 ppt 的海水中. 鰻魚孵化出來之後, 體型漸漸變成頭小身體透明扁平之柳葉狀, 故又稱柳葉鰻 (leptocephalus), 此體型構造爲外洋性浮游動植物之型態特徵之一. 此時柳葉鰻無自由行動之能力, 隨洋流漂浮至大陸邊緣, 依種類的不同, 此段時間預估約須一年以上, 在這段時間, 仔鰻之型態由柳葉狀變成長細型, 但仍爲透明故又稱玻璃鰻 (glass eel). 玻璃鰻已有自由行動之能力並對特定物質有驅近現象. 玻璃鰻在河口進入淡水後, 體表色素出現並長成爲小鰻. 由上述之鰻魚生活簡史, 可以知道鰻魚人工繁殖之困難點在: 之一, 影響鰻魚性成熟之內在 (例如: 內分泌, 體成份變化) 與環境 (例如: 鹽度, 溫度, 光照, 壓力…等等) 因子不明; 人工飼養之鰻魚或野生種鰻尚無在人爲環境下自然性成熟之紀錄. 之二, 仔鰻之孵化, 成長條件及初期餌料並不清楚, 若以一般海水魚類初期餌料, 如ss型海水輪蟲或戶外養成池池水, 最長僅可使幼苗存活一個月. 之三, 而目前鰻魚人工繁殖, 其受精卵之取得, 皆由長期注射異種生物激素而來, 使鰻魚處在不正常的生理狀況下; 報告指出, 既使在虹(魚尊)或鯉魚 使用人工催熟産生之幼苗, 比自然産卵受精孵化的幼苗, 具較高之死亡率和畸形率. 可見欲達成鰻魚人工繁殖之成功, 可能需由解決此三方面之問題齊下. 鰻魚的生殖生理的問題可能和其他魚類不同, 對水産養殖而言, 因爲其他魚種已經達性熟, 是欠缺引發堆積在腦下垂體中促性腺刺激素 (gonadotropins, GtHs) 的釋放 (discharge), 而引發配子的最後成熟. 所以所在早期水産養殖, 所謂的促性腺刺激素釋放激素的長效合成物 (gonadotropin-releasing hormone analogues, GnRH-a) 與多巴胺拮抗物 (anti-dopamine reagents) 有其應用上的價值. 而鰻魚的問題之一則是: 腦下垂體中的促性腺激素含量相當的低, 是故性腺刺激素釋放激素的長效合成物 (gonadotropin-releasing hormone analogues, GnRH-a) 或多巴胺拮抗物 (anti-dopamine reagents) 對鰻魚毫無效果的原因在此. 本文即在介紹有關鰻魚生殖內分泌的最新發展, 著重於腦下垂體中性腺刺激素生成的調控.
脊椎動物的生殖作用是由下視丘-腦下垂體-性腺所組成的神經內分泌軸線 (hypothalamus-pituitary-gonads neuroendocrine axis, HPG axis) 所控制. 腦部下視丘分泌的促性腺激素釋放激素 (gonadotropin-releasing hormone, GnRH) 控制腦下垂體中促性腺激素 (gonadotropins, 在高等脊椎動物是黃體激素, lutenizeing hormone, LH, 及濾泡刺激素, follicle-stimulating hormone, FSH; 在魚類則是促性腺刺激素一型, GtH-I, 及促性腺刺激素二型, GtH-II, 構造上 GtH-II 和 LH 較相近 而 GtH-I 則與 FSH 較相近, 其演化及生理意義以後再專文介紹之) 的生成和釋放; 而促性腺激素的作用則是刺激性腺的功能, 包括配子生成 (gametogenesis) 及類固醇激素的生成 (steroidogenesis). 在魚類, 促性腺激素的生成與釋放, 除了受促性腺激素釋激素 (GnRH) 的刺激外, 也同時由多巴胺 (dopamine) 所抑制. 而性腺所産生的各種激素 (如: 性類固醇激素), 能回饋控制腦部下視丘及腦下垂體, 進而影響達這條軸線的作用. 外部環境因子包括: 物理-化學, 營養及體內部狀況 (如: 生長及新陳代謝) 也能影響下視丘-腦下垂體-性腺神經內分泌軸的功能.
生物的發育並不是由稚體突然變成具生殖能力的成體, 而是需要 '青春期' 的過渡及準備. 所謂的 '青春期' (puberty) 是指生物稚體變成具有生殖能力成體的一段過程; 在這段過程中, 生物體會有某些改變, 包括: 外部型態, 生理狀況, 行爲…等等. 青春期的英文名辭 ‘puberty’ 是由 '體毛' (pubes) 這個拉丁字而來的, 指人類在青春期時某些部位的體毛(或體毛的生長). 後來泛指哺乳動物及其它脊椎動物, 包括: 鳥類及魚類的生殖發動準備期. 現在包括無脊椎動物, 都利用這個名辭.
在魚類, 青春期 (生殖發動準備期) 的認定, 在雄性, 爲精子 (spermatocyte) 的出現; 在雌性, 爲卵黃生成 (vitellogenesis) 的開始. 而青春期的完成則以第一次排卵或排精爲認定標準, 即個體具備生殖能力. 所以青春期是一個動態的演變現象, 在這一段時間內, 稚體完成具有生殖能力成體的所有準備. 從生殖內分泌的觀點來看, 最重要的是腦下垂體中的促性腺激素的生成和釋放, 因爲在所有的脊椎動物中, 促性腺激素對性腺的發育和配子的生成扮演重要的角色, 直接和生殖有關. 青春期的發動與個體的年齡及成長的體型有關, 因爲體型和體中堆積的能量有關, 而母體若有足夠的能量累積可, 則以確保母體生殖的成功和幼體發育的完整. 例如: 某些具生殖遷徙行爲的動物, 必須遷徙到適當環境, 才可能發生生殖行爲; 在卵生動物, 能量以卵黃由母體轉移到子代; 在哺乳動物, 母體能量經由胎盤及乳汁轉移到子代. 所以與生長及新陳代謝有關的激素, 可能在傳統的生殖激素外, 對生殖及青春期的啟動扮演重要的功能.
魚類和其他脊椎動物一樣, 已有有相當多的學術證據及現場經驗, 支持成長率和生殖啟動 (青春期) 的正相關性. 但到目前爲止, 卻只有很少的學術報告指明, 是何種化學訊息及生理機制連接體成長和青春期的啟動. 體成長主要受成表激素 (growth hormone, GH) 的調控, 但腦下垂體細胞培養實驗證明, 成長激素和促性腺激素的生成或釋放沒有直接的關係, 推測成長激素可能需要借由其他因子來連接體成長和生殖啟動. 最近的報告指出, 類胰島素成長因子 (insulin-like growth factors, IGFs) 在離體實驗條件下, 可促進鰻魚腦下垂體中促性腺激素的生成和釋放. 類胰島素成長因子的生成, 特別是類胰島素成長因子一型 (insulin-like growth factor-I, IGF-I), 受成長激素的控制, 主要的生成部在肝臟再釋放到血液循環中, 在其他組織中, 受成長激素的調控, 也有少量的區域性生成. 成長激素的作用需要類胰島素成長因子一型的協同作用, 才能發揮最大作用; 最明顯的例子, 魚類在饑餓狀態下, 爲調節能量平衡, 血液中有高量的成長激素, 可是此時成長卻是被抑制的, 同時類胰島素成長因子一型的血液含量也是很低. 而其他和生長及發育有關的激素, 例如甲狀腺素 (thyroxins) 已被證明對鰻魚腦下垂體中促性腺激素的生成無任何作用. 顯示類胰島素成長因子爲連結體成長和生殖啟動的化學訊息. 不止在鰻魚, 最近的報告也指出, 在鮭鱒魚類中, 類胰島素成長因子一型也可以刺激腦下垂體中促性腺激素的含量, 同時也能加強促性腺激素釋放激素對刺激腦下垂體性腺刺激素釋放的效能. 這一點在哺乳動中也有相同的結果. 以上的證據說明, 除了刺激體成長外, 類胰島素成長因子一型也可以直接刺激素腦下垂體中促性腺刺激素的生成和釋放, 更確定類胰島素成長因子一型是連接體成長和青春期 (生殖) 啟動的化學因數.
所謂良好的生殖體型包括: 體型大小及足夠的能量累積, 以應付在生殖過程中所需的能量. 和其他脊椎動物一樣, 胰島素 (insulin) 在魚類中, 對蛋白質和脂肪質的同化代謝作用扮演著極重要的功能. 在鮭魚的銀化 (smoltification) 和兩棲類的變態, 胰島素被證明具有觸發啟始的作用, 可能與其對能量轉換所具有的重要影響有關. 在鰻魚, 實驗證明胰島素的注射可顯著的提高鰻魚腦下垂體中促性腺激素的含量. 對鰻魚而言, 銀鰻 (成鰻) 比黃鰻 (小鰻) 體成份中含更高量的脂肪. 這種現象使學者提出脂質的堆積到某一個臨界量, 是推動鰻魚進入生殖發動期的重要因素, 這種概念在哺乳動物中早已經被提出. 在哺乳動物中, 目前能量堆積和生殖作用彼此交互影響的研究方面, 抑脂激素 (leptin) 是一個重點, 抑脂激素是由脂肪細胞所製造分泌, 它和食欲及攝食行爲有關. 目前有相當多的報告指出, 抑脂激素和促性腺激素釋放激素的作用有關, 而肥胖症和某些營養不良引起的不孕, 可用由抑脂素的處理而得到矯正. 雖然抑脂激素在魚類中的存在尚未被確定, 但是由分子生物技術確定在鰻魚中有此基因的存在. 同時, 銀鰻的體脂肪含量可高達 30% 以上, 抑脂激素的存在可能對鰻魚的生殖具有某些功能, 而離體腦下垂體細胞培養的實驗證明, 抑脂激素有促進鰻魚促性腺激素含量的功效, 同時也可刺激虹鱒促性腺激素的釋放. 由這些證據的支援了, 在魚類中體脂肪質的堆積和生殖發動的生理基礎, 可能是藉由胰島素和抑脂激素所傳遞的.
在青春期中, 特別是卵生動物, 身體所貯藏的能量必須轉移到性腺的發育 (包括配子細胞及性腺細胞本身的發育). 某些魚類, 例如鰻魚或鮭魚在生殖期時呈斷食的狀態而且需要進行長達數千公里的生殖回游, 這些能量均需在青春期中準備堆積好. 以鰻魚爲例, 在銀鰻階段時即開始生理性斷食, 而性成熟的雌魚其性腺體重比 (GSI) 可由原來的 2 % 成長到 50 % 以上. 而這現象是在鰻魚回游數千公里後發生的, 身體所貯藏的能量機乎完全轉移到性腺的發育. 可體素 (cortisol) 由其腎上腺皮質所製造分泌, 對生物的能量中間代謝有關包括屯積脂肪的運用, 及刺激醣質新生作用 (gluconeogenesis), 些作用在脊椎動物中是共通的. 在哺乳動物中, 可體素除了被證明和壓力的有關, 會抑制生殖作用之外, 同時也被證明和青春的啟動有關, 在魚類也有相同的報告, 抑制成魚生殖作用, 卻刺激稚魚, 包括虹鱒及鰻魚, 腦下垂體促性腺素的含量. 對鰻魚而言,活體和離體細胞腦下垂體細胞培養都證明了可體素具有刺激素腦下垂體促性腺素生成作用. 這些證據顯示可體素對生殖的作用, 除了在能量中間代謝層面外, 也可直接作用在腦下垂體刺激性腺刺激素的生成.
由上述的實驗中的結果說明, 魚類的腦下垂體中的促性腺激素細胞, 除了受傳統的下視丘-腦下垂體-性腺所組成的神經內分泌軸線所控制, 包括: 促性腺激素釋放激素, 多巴胺, 促性腺激素及性類固醇的調控之外, 某些所謂週邊訊息 (peripheral messengers) , 包括: 體成長, 新陳代謝情形, 組織貯能狀況, 可藉由如類成胰島素成長因子 (IGFs), 可體素 (cortisol), 胰島素 (insulin) 及抑脂激素 (leptin) 等而達到生殖調控的目的.
事實上大家早就了解種魚選擇的標準之一, 就是肥滿度, 包括體型及觸感所得的性腺發育情形. 而鰻魚, 特別是養殖鰻, 從生殖生理的觀點並無所謂的 '種鰻', 只有長得比較大的鰻.謹以此文拋磚引玉, 歡迎大家一起討論.
主要參考文獻:
- Dufour S., Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Vidal B., Marchelidon J., Querat B., Burzawa-Gerard E., Chang C-F., Schmitz M. (1999) Puberty in teleosts: new insights into the role of peripheral signals in the stimulation on pituitary gonadotropins. Proceedings of thr 6th International Symposium on the Reproductive Physiology of Fish, Bergen, Norway, July 4-9, 1999. (in press).
- Huang Y-S., Rousseau K., Le Belle N., Vidal B., Burzawa-Gerard E., Marchelidon J. and Dufour S. (1998) Insulin-like growth factor I (IGF-I) stimulates gonadotropin production from eel pituitary cells: a possible metabolic signal for induction of puberty. J. Endocrinol. 159:43-52.
- Huang Y-S. (1998) Role des steroides sexuels et des hormones metaboliques dans le controle direct hypophysaire de l’hormone gonadotrope (GtH-II) chez l’anguille europeenne, Anguilla anguilla. Universite Paris VI, Departement de Physiologie de la Reproduction, 173p.
- Huang Y-S., Rousseau K., Sbaihi M., Le Belle N., Schmitz M. and Dufour S. (1999) Cortisol selectively stimulates pituitary gonadotropin (GtH-II) subunit, in a primitive teleost, Anguilla anguilla. Endocrinology 140:1228-1235.
- Huang Y-S., Rousseau K., Le Belle N., Vidal B., Burzawa-Gerard E., Marchelidon J. and Dufour S. (1999) Opposite effects of insulin-like growth factors (IGFs) on gonadotropin (GtH-II) and growth hormone (GH) production by primary culture of European eel ( Anguilla anguilla ) pituitary cells. Aquaculture 177:73-83.
-Rousseau K., Huang Y-S., Le Belle N., Vidal B., Marchelidon J., Epelbaum J. and Dufour S. (1998) Long-term growth hormone release by serum-free primary culture of pituitary cells from European eel ( Anguilla anguilla ): inhibitory effects of somatostatin and insulin-like growth factors. Neuroendocrinology 67:301-309.
故事(二)
關於鰻魚的身世之謎,突然想到世說新語上的一個故事:謝太傅寒雪日內集,與兒女講論文義。俄而雪驟,公欣然曰:「白雪紛紛何所似?」兄子胡兒曰:「撒鹽空中差可擬。」,兄女曰:「未若柳絮因風起。」公大笑樂。 翻成白話文就是:宋朝謝安有一天和晚輩上國文課,突然下雪,謝安很高興的說:「下雪像什麼」 ? 他的姪子搶道:「和向天灑鹽巴差不多」 。姪女比較細膩,說:「倒不如是柳絮被風吹起!」 ,謝安聽了大樂而笑。 要是我,我也會說 :「撒鹽空中差可擬。」因為20幾年前在雪山 369 山莊看到的是如灑鹽般的下雪 (後來在畢里牛斯山看到的是鵝毛) 。關於追蹤鰻魚產卵地,我想不到好的形容詞,姑且稱為「大海撈針」。
上次提到,經過兩千多年才確定鰻魚也是有父有母的,但還沒人知道他們在哪裡交配產卵。事實上,西方人把鰻魚生活史稱為奧迪賽 (odysseys就是起源取自古羅馬希臘神話奧迪賽,就是必須離開自己王國,出外去做一趟危險的旅程) 般的史詩。
再複習鰻魚特殊的生活史,在淡水環境中生長數年後(雄魚約4-5 年,雌魚約6-7 年以上),則須降海進行生殖,如果不降海則永遠無法達到自然性成熟。鰻魚降海之後,歐洲鰻在海洋洄游數4千多公里後在藻海(Sargasso Sea) 附近 達深達百米以上的產卵場。日本鰻則游了6千多公里在馬里亞那群島 (Mariana Islands) 附近產卵場。鰻魚孵化出來之後, 體型漸漸變成頭小身體透明扁平, (leptocephalus),故其拉丁文之意義為”狹(lepto-)” “頭(cephal-)” 魚,此體型構造爲外洋性浮游動植物之型態特徵之一,最早,人們並不知道柳葉鰻是鰻魚的幼苗。柳葉鰻無自由行動之能力,隨洋流漂浮至大陸邊緣,此段時間預估約須一年以上,在這段時間,仔鰻之型態由柳葉狀變成長細型, 但仍爲透明故又稱玻璃鰻(glass eel),此為第一次變態,玻璃鰻已有自由行動之能力,並對特定物質有驅近現象。玻璃鰻在進入淡水後,體表色素出現成為小鰻。小鰻(elver)再長成黃鰻 (yellow eel),黃鰻在淡水中成長數至十數年變態成銀鰻 (silver eel),此為第二次變態。
故事的開始是:19世紀有人在地中海西西里島東北方的 Messina 海峽捉到 一尾怪魚,體型頭小身體透明扁平,在1856年被 Kaup命為狹頭魚 (leptocephalus) 取其拉丁文之意義為”狹(lepto-)” “頭(cephal-)”,很可惜的是 這101 條後來遺失了 (好像跟北京人的頭骨命運一樣…),幸好四十幾年後,義大利有兩個好事者 叫 Grassi 與 Calandruccio 在同一個地方捉了許多狹頭魚,從1887 到 1892年放在水族缸中蓄養 (這八成是西方世界養鰻的濫觴….) 。結果在1896年竟發現狹頭魚變成淡水鰻魚,終於知道鰻魚會變態,而且是從海裡來的…
下一步就是找尋鰻魚的產卵地。從地圖上來看,西西里島位於地中海中,而地中海以直布駝羅海峽(Gibraltar) 和大西洋相接,而且經過調查,南歐、地中海、西北歐大西洋岸、英國沿岸、英倫海峽 (法國人叫 La Manche,英國人叫 English Channel ) 所捉到的小鰻魚(玻璃鰻) 體型和型態多差不多,所以他們認為,鰻魚應該是在離岸不遠處產卵。可惜,沿岸漁業一直沒捉到大腹便便的母鰻不說,連比狹頭魚更小的魚都捉不到。事件似乎又陷入另一個迷雲中….。
就在此時 (1905年) 丹麥政府派遣一位單丹麥籍船長叫 Johannes Schmidt去尋找鰻魚的產卵地,在此之前Schmidt曾成功的找到鱈魚的產卵場。他在北大西洋的正中心 Faores 群島西邊捉到狹頭魚,這是第一次在地中海外捉到狹頭魚,意義重大 (諸位看倌 應該聽過 大海撈針是用來形容 mission impossible 不過捉狹頭魚 真的就是大海撈針 其體長不過4-1公分 而且是透明的…)。所以從 1905 到1920 年 Schmidt 就在大西洋上來來回回的進行大海撈針。其間,他的船跡北到冰島,南到迦納利群島 (Canary islands),東橫渡地中海到埃及 西邊到美洲大陸,(好像比西遊記的14年還累...),不斷的海底撈針,並參考洋流,比對抓到魚的型態大小(體長、脊椎骨數、鰭條數)。從1908年到1910年,他發現在地中海中,愈往直布駝羅海峽西邊捉到的愈大(即愈深入地中海東邊)。所以他認為鰻魚應該在大西洋中產卵。剛好191年時,Hjort 在Azores群島附近捉到一些 4-6公分的小魚,是有紀錄以來的最小體型,所以 Hiort認為鰻魚的產卵場應位於Azores群島和百慕達群島之間。1911-1915年共家入23條船在550個採集點,共捉到120條狹頭鰻 (不過 1914-1918 第一次世界大戰影響他的調查,部分調查船被德國的潛艇無限戰擊沉,他只好回陸地 研究狹頭魚型態分類)。終於在1920年於大西洋西側靠近北美洲外海,他開著四桅動力帆船,DANA 號,在藻海處 (Sargasso Sea, 馬尾藻海或稱為薩加索, 葡語葡萄果的意思 因為藻體由固養器、主幹、藻葉和氣囊四部分構成,而氣囊長得像葡萄,故稱 大致。在北緯20~35°、西經35~70°之間,覆蓋大約500~600萬平方公裡的水域) 捉到1公分長的小魚,此後未曾有比此小的體型被捉的紀錄,所以產卵場應該在這附近。但很可惜,沒有發現卵。附帶一點 Johannes Schmidt 在調查歐洲鰻的過程中連帶的也把美洲鰻的產卵場釐清,發現也是在藻海附近,只不過比較偏西一點。 經過16 年的努力,終於解開持續2千多年的迷團。
1956年時Leon Bertin.於他的書 (Eel: A Biological Study) 指出:「鰻魚在藻海產卵,這沒有錯,但更精確的講,她們只在海藻漂浮面中心的深處約400米,溫度16-17度的地方產卵...」。 很有趣的是,雖然美洲鰻和歐洲鰻產卵地點很近,不過,在北美洲找不到歐洲鰻 在歐洲找不到美洲鰻。後來有好事者,因為藻海離百慕達三角很近, 又八卦認為此區是外星人出沒之地,。故鰻魚是外星生物…。又有一說,據說古代在大西洋中有一塊大陸名為亞特蘭提斯,鰻魚就是回到那裡去產卵。
而在亞洲這邊,特別是東北亞,日本學者早在 1930 年代就開始有「大海撈針」的報告 ( Matsui, 1957)。 他們根據鰻魚的地理分布與海流輸送的關聯,推定產卵場應該是在台灣以東,琉球以南的海域。 所以日本人就埋頭在這海域進行調查。可惜30年過了,卻毫無結果。 在1986年夏天,日本東京大學海洋研究所的研究船在菲律賓東北海域進行調查,捉到21尾狹頭魚,最小為3.39公分,加上北赤道洋流, 斷定日本鰻應該是在馬里亞那海溝附近產卵 (Kajihara, 1988)。最後在 1991年7月,日本東京大學海洋研究所的研究船「白鳳丸 (Hakuho Maru) 」,在 塚本勝己 (Katsumi Tsukamoto) 副教授的帶隊下,一口氣破天荒捉到 911 條狹頭魚,這比 日本 30 幾年來所捉的總數還要多。其中在北緯 15 度 東經 137 度所捉到的 239 條中, 最小的只有 0.79 公分,預估是孵化後 10 天的苗,證實日本鰻是在馬里亞那群島西側產卵 (Tsukamoto, 1992),日本人就把成果發表在 Nature 雜誌上,結果那一期的Nature (1992, No.356)封面就是以捉到的狹頭魚當封面主角 (很有趣的是,群體作業但整篇報告只有一位作者, 和現在掛一大串作者大相逕庭)。
台灣呢 ? 號稱養鰻大國,當然在這方面不能留空白。在1995年8月16-23日,台灣省水產試驗所「水試一號」試驗船在菲律賓以東的海域,捕獲三尾是日本鰻。這是台灣有史以來,第一次捕獲日本鰻的柳葉形仔鰻。三尾柳葉形仔鰻的體長分別為 2.74,2.88 以及 3.14 公分,其日齡分別為46,50及51天。本結果支持日本鰻是在夏季產卵的推論,但根據採集地點與海流方向,這三尾柳葉形仔鰻之體長與日齡 水是所人員認為 不足以肯定或說明目前所認知的產卵埸,而推定其產卵場之分布範圍應相當廣闊。此後隨著省水產試驗所的改隸與組織、政策的調整,這一頁恐被埋入歷史中。中國大陸則是在1996 年聲稱於1993年3月海測船「向陽紅05 號」在產卵場附近採到19粒鰻魚卵 !?日本比較保守,宣稱撈到三個。不過後來利用遺傳標記又推翻那三個長得很像的魚卵…。
雖然,歸納天文、洋流、氣候、小魚型態與生態之特性、並配合耳石 (otolith 等於魚的黑盒子 不過不是通話紀錄器 而是飛行數據紀錄器)日週輪而找出歐洲鰻與日本鰻產卵場,但迷團還是沒被解,至少沒親眼看到大腹便便的親魚或撈到受精魚卵。但漸漸的,幾個結論被歸納出來 1. 開發現鰻產卵場和海山(sea mountain, 海下山峰) 有關; 2.交配產卵與月週期有關, 鰻苗在 5, 6月新月(New moon)時孵化, 稱為新月假說 (new moon hypothesis); 3.歐洲鰻或日本鰻之產卵分別與溫度鋒 (thermal front)或 鹽度鋒 (salinity front)有關 (即溫度或鹽度變化的介面)。
To see is to believe, 所以 1998年日本和德國 (瞧!軸心國又來了, 50幾年前應該就只合搞這個就好了..) 在生殖季節時乾脆派潛艇下海去 (日本派船,德國派潛艇叫 「JAGO號」, 性能諸元請查http://www.mpi-seewiesen.mpg.de/~hissmann/jago.html) 調查。」JAGO 號」可搭乘2人,潛航深度 400 米。他們在馬里亞那群島附近的 Arakane 與 Pathfinder 海山,在新月前後進行91 小時的調查。結果, 鰻魚還是有廉恥心的,燕好時不會大肆張揚。所以偷拍不成,他們便下推論:肯定在深於 400 米以下產卵… 。故事還是會繼續的…。
以追蹤小魚配合海流、耳石尋找產卵地,反過來說倒不如叫識途老馬帶路:把牠們植入晶片或追蹤器再去找。可惜,外星人不出來認養他的寵物,不然 CIA, FBI, NASA 一定出面,像電影全民公敵 (Enemy of the State) 或 X 檔案 (X-Files)。 因為目前用的發報器、追蹤器功率太小,只知道鰻魚下海後會做 上下 200 公尺的游動,約3-4天後就潛到追蹤不到的深度。人類在登陸火星之前,不知道能不能解開這個謎?
至於鰻魚研究、鰻魚人工繁殖下回分解
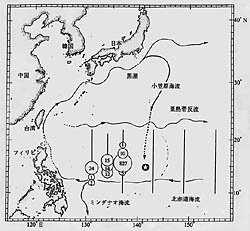

圖1. 歐洲鰻與日本鰻之產卵場。 (摘自 The European Freshwater Eel, 1975; 及 Nature, 1992)。

JAGO 潛艇,隸屬於德國 「馬克司-蒲朗克研究所 (Max Planck Institute)』。

Nature (1992, No.356) 的封面。

「大海撈針」的工具
上次提到,經過兩千多年才確定鰻魚也是有父有母的,但還沒人知道他們在哪裡交配產卵。事實上,西方人把鰻魚生活史稱為奧迪賽 (odysseys就是起源取自古羅馬希臘神話奧迪賽,就是必須離開自己王國,出外去做一趟危險的旅程) 般的史詩。
再複習鰻魚特殊的生活史,在淡水環境中生長數年後(雄魚約4-5 年,雌魚約6-7 年以上),則須降海進行生殖,如果不降海則永遠無法達到自然性成熟。鰻魚降海之後,歐洲鰻在海洋洄游數4千多公里後在藻海(Sargasso Sea) 附近 達深達百米以上的產卵場。日本鰻則游了6千多公里在馬里亞那群島 (Mariana Islands) 附近產卵場。鰻魚孵化出來之後, 體型漸漸變成頭小身體透明扁平, (leptocephalus),故其拉丁文之意義為”狹(lepto-)” “頭(cephal-)” 魚,此體型構造爲外洋性浮游動植物之型態特徵之一,最早,人們並不知道柳葉鰻是鰻魚的幼苗。柳葉鰻無自由行動之能力,隨洋流漂浮至大陸邊緣,此段時間預估約須一年以上,在這段時間,仔鰻之型態由柳葉狀變成長細型, 但仍爲透明故又稱玻璃鰻(glass eel),此為第一次變態,玻璃鰻已有自由行動之能力,並對特定物質有驅近現象。玻璃鰻在進入淡水後,體表色素出現成為小鰻。小鰻(elver)再長成黃鰻 (yellow eel),黃鰻在淡水中成長數至十數年變態成銀鰻 (silver eel),此為第二次變態。
故事的開始是:19世紀有人在地中海西西里島東北方的 Messina 海峽捉到 一尾怪魚,體型頭小身體透明扁平,在1856年被 Kaup命為狹頭魚 (leptocephalus) 取其拉丁文之意義為”狹(lepto-)” “頭(cephal-)”,很可惜的是 這101 條後來遺失了 (好像跟北京人的頭骨命運一樣…),幸好四十幾年後,義大利有兩個好事者 叫 Grassi 與 Calandruccio 在同一個地方捉了許多狹頭魚,從1887 到 1892年放在水族缸中蓄養 (這八成是西方世界養鰻的濫觴….) 。結果在1896年竟發現狹頭魚變成淡水鰻魚,終於知道鰻魚會變態,而且是從海裡來的…
下一步就是找尋鰻魚的產卵地。從地圖上來看,西西里島位於地中海中,而地中海以直布駝羅海峽(Gibraltar) 和大西洋相接,而且經過調查,南歐、地中海、西北歐大西洋岸、英國沿岸、英倫海峽 (法國人叫 La Manche,英國人叫 English Channel ) 所捉到的小鰻魚(玻璃鰻) 體型和型態多差不多,所以他們認為,鰻魚應該是在離岸不遠處產卵。可惜,沿岸漁業一直沒捉到大腹便便的母鰻不說,連比狹頭魚更小的魚都捉不到。事件似乎又陷入另一個迷雲中….。
就在此時 (1905年) 丹麥政府派遣一位單丹麥籍船長叫 Johannes Schmidt去尋找鰻魚的產卵地,在此之前Schmidt曾成功的找到鱈魚的產卵場。他在北大西洋的正中心 Faores 群島西邊捉到狹頭魚,這是第一次在地中海外捉到狹頭魚,意義重大 (諸位看倌 應該聽過 大海撈針是用來形容 mission impossible 不過捉狹頭魚 真的就是大海撈針 其體長不過4-1公分 而且是透明的…)。所以從 1905 到1920 年 Schmidt 就在大西洋上來來回回的進行大海撈針。其間,他的船跡北到冰島,南到迦納利群島 (Canary islands),東橫渡地中海到埃及 西邊到美洲大陸,(好像比西遊記的14年還累...),不斷的海底撈針,並參考洋流,比對抓到魚的型態大小(體長、脊椎骨數、鰭條數)。從1908年到1910年,他發現在地中海中,愈往直布駝羅海峽西邊捉到的愈大(即愈深入地中海東邊)。所以他認為鰻魚應該在大西洋中產卵。剛好191年時,Hjort 在Azores群島附近捉到一些 4-6公分的小魚,是有紀錄以來的最小體型,所以 Hiort認為鰻魚的產卵場應位於Azores群島和百慕達群島之間。1911-1915年共家入23條船在550個採集點,共捉到120條狹頭鰻 (不過 1914-1918 第一次世界大戰影響他的調查,部分調查船被德國的潛艇無限戰擊沉,他只好回陸地 研究狹頭魚型態分類)。終於在1920年於大西洋西側靠近北美洲外海,他開著四桅動力帆船,DANA 號,在藻海處 (Sargasso Sea, 馬尾藻海或稱為薩加索, 葡語葡萄果的意思 因為藻體由固養器、主幹、藻葉和氣囊四部分構成,而氣囊長得像葡萄,故稱 大致。在北緯20~35°、西經35~70°之間,覆蓋大約500~600萬平方公裡的水域) 捉到1公分長的小魚,此後未曾有比此小的體型被捉的紀錄,所以產卵場應該在這附近。但很可惜,沒有發現卵。附帶一點 Johannes Schmidt 在調查歐洲鰻的過程中連帶的也把美洲鰻的產卵場釐清,發現也是在藻海附近,只不過比較偏西一點。 經過16 年的努力,終於解開持續2千多年的迷團。
1956年時Leon Bertin.於他的書 (Eel: A Biological Study) 指出:「鰻魚在藻海產卵,這沒有錯,但更精確的講,她們只在海藻漂浮面中心的深處約400米,溫度16-17度的地方產卵...」。 很有趣的是,雖然美洲鰻和歐洲鰻產卵地點很近,不過,在北美洲找不到歐洲鰻 在歐洲找不到美洲鰻。後來有好事者,因為藻海離百慕達三角很近, 又八卦認為此區是外星人出沒之地,。故鰻魚是外星生物…。又有一說,據說古代在大西洋中有一塊大陸名為亞特蘭提斯,鰻魚就是回到那裡去產卵。
而在亞洲這邊,特別是東北亞,日本學者早在 1930 年代就開始有「大海撈針」的報告 ( Matsui, 1957)。 他們根據鰻魚的地理分布與海流輸送的關聯,推定產卵場應該是在台灣以東,琉球以南的海域。 所以日本人就埋頭在這海域進行調查。可惜30年過了,卻毫無結果。 在1986年夏天,日本東京大學海洋研究所的研究船在菲律賓東北海域進行調查,捉到21尾狹頭魚,最小為3.39公分,加上北赤道洋流, 斷定日本鰻應該是在馬里亞那海溝附近產卵 (Kajihara, 1988)。最後在 1991年7月,日本東京大學海洋研究所的研究船「白鳳丸 (Hakuho Maru) 」,在 塚本勝己 (Katsumi Tsukamoto) 副教授的帶隊下,一口氣破天荒捉到 911 條狹頭魚,這比 日本 30 幾年來所捉的總數還要多。其中在北緯 15 度 東經 137 度所捉到的 239 條中, 最小的只有 0.79 公分,預估是孵化後 10 天的苗,證實日本鰻是在馬里亞那群島西側產卵 (Tsukamoto, 1992),日本人就把成果發表在 Nature 雜誌上,結果那一期的Nature (1992, No.356)封面就是以捉到的狹頭魚當封面主角 (很有趣的是,群體作業但整篇報告只有一位作者, 和現在掛一大串作者大相逕庭)。
台灣呢 ? 號稱養鰻大國,當然在這方面不能留空白。在1995年8月16-23日,台灣省水產試驗所「水試一號」試驗船在菲律賓以東的海域,捕獲三尾是日本鰻。這是台灣有史以來,第一次捕獲日本鰻的柳葉形仔鰻。三尾柳葉形仔鰻的體長分別為 2.74,2.88 以及 3.14 公分,其日齡分別為46,50及51天。本結果支持日本鰻是在夏季產卵的推論,但根據採集地點與海流方向,這三尾柳葉形仔鰻之體長與日齡 水是所人員認為 不足以肯定或說明目前所認知的產卵埸,而推定其產卵場之分布範圍應相當廣闊。此後隨著省水產試驗所的改隸與組織、政策的調整,這一頁恐被埋入歷史中。中國大陸則是在1996 年聲稱於1993年3月海測船「向陽紅05 號」在產卵場附近採到19粒鰻魚卵 !?日本比較保守,宣稱撈到三個。不過後來利用遺傳標記又推翻那三個長得很像的魚卵…。
雖然,歸納天文、洋流、氣候、小魚型態與生態之特性、並配合耳石 (otolith 等於魚的黑盒子 不過不是通話紀錄器 而是飛行數據紀錄器)日週輪而找出歐洲鰻與日本鰻產卵場,但迷團還是沒被解,至少沒親眼看到大腹便便的親魚或撈到受精魚卵。但漸漸的,幾個結論被歸納出來 1. 開發現鰻產卵場和海山(sea mountain, 海下山峰) 有關; 2.交配產卵與月週期有關, 鰻苗在 5, 6月新月(New moon)時孵化, 稱為新月假說 (new moon hypothesis); 3.歐洲鰻或日本鰻之產卵分別與溫度鋒 (thermal front)或 鹽度鋒 (salinity front)有關 (即溫度或鹽度變化的介面)。
To see is to believe, 所以 1998年日本和德國 (瞧!軸心國又來了, 50幾年前應該就只合搞這個就好了..) 在生殖季節時乾脆派潛艇下海去 (日本派船,德國派潛艇叫 「JAGO號」, 性能諸元請查http://www.mpi-seewiesen.mpg.de/~hissmann/jago.html) 調查。」JAGO 號」可搭乘2人,潛航深度 400 米。他們在馬里亞那群島附近的 Arakane 與 Pathfinder 海山,在新月前後進行91 小時的調查。結果, 鰻魚還是有廉恥心的,燕好時不會大肆張揚。所以偷拍不成,他們便下推論:肯定在深於 400 米以下產卵… 。故事還是會繼續的…。
以追蹤小魚配合海流、耳石尋找產卵地,反過來說倒不如叫識途老馬帶路:把牠們植入晶片或追蹤器再去找。可惜,外星人不出來認養他的寵物,不然 CIA, FBI, NASA 一定出面,像電影全民公敵 (Enemy of the State) 或 X 檔案 (X-Files)。 因為目前用的發報器、追蹤器功率太小,只知道鰻魚下海後會做 上下 200 公尺的游動,約3-4天後就潛到追蹤不到的深度。人類在登陸火星之前,不知道能不能解開這個謎?
至於鰻魚研究、鰻魚人工繁殖下回分解


圖1. 歐洲鰻與日本鰻之產卵場。 (摘自 The European Freshwater Eel, 1975; 及 Nature, 1992)。

JAGO 潛艇,隸屬於德國 「馬克司-蒲朗克研究所 (Max Planck Institute)』。

Nature (1992, No.356) 的封面。

「大海撈針」的工具
故事

鰻魚屬於鰻鱺科(Anguillidae),鰻屬(Anguillus),是一種非常特殊的魚類,全世界總共有十八種。鰻魚因為奇特的生活史,一直以來對鰻魚的生態都是個謎,因此全世界對其研究從未間斷過;台灣地區鰻魚的種類有四種,最常見者為日本鰻( Anguilla japonica) 及鱸鰻(A. marmorata);但是西里伯斯鰻(A. celebesensis) 及短鰭鰻(A. bicolor pacifica) 則比較罕見。鰻魚在演化上屬於非常原始的低等脊椎動物,其生長到不同時期時會有變態發生而改變外觀;跟鮭魚有著相反的生活史,鰻魚會在河川中長大、成熟後洄游到海洋中產卵,一生只產一次卵,產卵之後就死亡。鰻魚在卵孵化之後,最先形成柳葉鰻(Letocephalus)、然後變態成玻璃鰻(glass eel)、再來是鰻線(elvers)、黃鰻(yellow eel) ,最後變態成銀鰻(silver eel) 而降海產卵。
鰻魚(Anguilla spp.)具特殊的生活史,在淡水環境中生長數年後(雄魚約4-5 年 雌魚約6-7 年以上),則須降海進行生殖。雖然曾有養殖37年的非正式紀錄及體型重達5.1公斤、體長130公分的野生鰻被捕獲的記載,但如果不降海則永遠無法達到自然性成熟。鰻魚降海之後,在海洋洄游數千公里後到達深達百米以上的產卵場。歐洲鰻的產卵場推測在藻海附近,日本鰻的產卵場推測在馬里亞那群島附近。日本學者曾預測鰻魚在水深75-100米左右的水深孵化,由人工繁殖之經驗推測,鰻魚之受精卵應爲浮性卵,卵徑約在 1mm左右,可漂浮在 34 ppt 的海水中。鰻魚孵化出來之後, 體型漸漸變成頭小身體透明扁平,又稱柳葉鰻 (leptocephalus),故其拉丁文之意義為”狹(lepto-)” “頭(cephal-)” 魚,此體型構造爲外洋性浮游動植物之型態特徵之一,最早,人們並不知道柳葉鰻是鰻魚的幼苗。柳葉鰻無自由行動之能力,隨洋流漂浮至大陸邊緣,此段時間預估約須一年以上,在這段時間,仔鰻之型態由柳葉狀變成長細型, 但仍爲透明故又稱玻璃鰻(glass eel),此為第一次變態,玻璃鰻已有自由行動之能力,並對特定物質有驅近現象。玻璃鰻在進入淡水後,體表色素出現成為小鰻。小鰻(elver)再長成黃鰻 (yellow eel),黃鰻在淡水中成長數至十數年變態成銀鰻 (silver eel),此為第二次變態。
事實上,鰻魚之生活史,目前已經知道部份,但未完全清楚。稍微考究一下,這幾乎是耗時2千年,遍及中西的事件。
最早西方人認為,馬尾巴若掉到水裡會變成鰻魚;但早在希臘時代,亞里斯多德(Aristotle,自然史的作者,大概是最早的生物教科書出版商) 已經注意到,從沒看到過大肚子的鰻魚,也沒有看到鰻魚交配或生蛋,故阿德就認為水中的雜物自然生出鰻魚,就像破布、雜物堆在一起會生出老鼠一樣,此為「無生源論」(Abiogenesis)之理論由來,因為交白卷總是不能增加篇幅的…。但阿德至少注意到某一段時間鰻魚會向下游方向游去,而且也會游回來。後來羅馬時代的動物學家布利尼 (Pliny,動物誌的作者),認為鰻魚怎麼可能是「無生源論」(文人相輕,自古而然…),他認為鰻魚用自己身體來摩擦岩石,掉下來的碎片就生成小鰻魚(瞧!雷爾教派的祖師爺,所以義大利人骨子裡有無性複製的思想血統)。一直到13世紀,德國哲學家Albertus Magnus還支持此理論。到16世紀,魚類分類的祖師爺 Rondelet (於1554年出版魚類全誌)還認為鰻魚是由腐壞的東西生出來的 (好像小時後,我媽也是這樣說我是從垃圾堆撿來的一樣),但他承認蛋是由雌雄交配後產生的…。後來雷文霍克(Leeuwenhoek, 發明顯微鏡的那一個人) 與林奈(Linnaeus, 發明二名法來命名物種的那個人),因為找到卵胎生蝦虎魚(blenny或稱尉魚),此類魚體內授精,魚卵在體內孵化,等生出母體就是一條不折不扣的小魚,好巧不巧,雷文霍克在鰻魚魚螵內找到寄生線蟲,所以他認定魚螵是鰻魚的子宮,而寄生線蟲是幼鰻 (事實上,螵線蟲真的長的和鰻魚一樣,只是小很多),這個錯誤一直到17世紀才被另一位義大利Redi (應該是弗羅倫斯,也就是徐志摩說的斐冷翠,因為彼時義大利還未出現)的學者更正它是寄生線蟲,而鰻魚的卵巢首次被討論記載,但因為鰻魚卵巢實在太不成熟,和一般認知的不同,所以並沒有被全面接受;Redi 並提出鰻魚的產卵地在海裏的假說 (不知道有沒有引起騷動,先知是寂寞的…)。一直到1850年,德國動物學家 Rathke (提出“重演律”-個體胚胎發生是種系發生的重演之人)詳細的描述其生殖腺,從此以後,鰻魚的卵巢才被確定。很好玩的是,雄鰻的精巢,要在24年後的1874年,才被一位波蘭學者(Syrski, 其任職於博物館)找到,為什麼?賣個關子,如果你已經耐心的讀到這裡,想一想,下回分曉。從此以後幼鰻的精巢又叫 Syrski organ。剛好在同一年,另一位德國的學者 (Munter, 任職於格賴夫斯瓦爾德的博物館) 在檢視了3000 條鰻魚後,竟然沒找到半條公的 (我現在做實驗剛好相反,養殖鰻很難找到一條母的),所以’罪證確鑿’,他說鰻魚是孤雌生殖(parthenogenetic)….。終於,在搞了兩千多年後,最後確定鰻魚不是怪胎,非石頭蹦出、垃圾桶揀的。
至於東方的傳說呢?反正有山就有水,有水就有魚,有魚大家摸,中國人一向是樂天知命,(正確的來說,應是肚子多填不飽,看到魚先想到紅燒或清燉,哪管從哪裡來;鰻非我族類,究其源?此非經世濟國之道也,故大丈夫不為也,省得落入玩物喪志,列入文化基本教材,禍遺子孫)。找得到文獻記載的,首推清康熙的古今圖書集成 (相當於大清百科全書) ; 鰻:一作鰻鱺,一作白鱔。埤雅云:「無鱗甲,白腹似鱔而大,青色;焚其骨,煙氣闢蠹,有雄無雌,以影漫鱧」。趙辟公雜說云:「有鰻鱺音,以影漫於鱧魚,擇其子接附鱧之鬐鬣而生,故謂之鰻鱺也。」。
台灣通志:「鰻,有烏白兩種,烏者為上,有雄無雌,以影鰻鱧生子而諺云:「鰻出鱧魚背」。相傳:鰻子在醴背上,如刳鰻有子者,必毒。又鱔腹無子者,亦毒,均當棄之」 (文獻摘自http://webpac.nhu.edu.tw/tr/)。
曰何?試解如後:我們還是比較講究實(食)用主義的,鰻魚又叫鰻鱺或白鱔,它沒有鱗片(非也它還是有鱗片的,鱗片細小,長橢園形,退化埋於皮下,故曰無鱗),肚子是白色的,背部是青色的(此樣本鑑定為”黃鰻”),把它的骨頭拿來燒,可以趨趕書蟲(生物防治法?),只有雄魚無雌魚 (和1850年德國動物學家Rathke結論不同,其中大有學問,這個牽涉到東西方治學的方法),身影長得和烏鱧魚差不多(好像差蠻多的?烏鱧魚即現今的魚種”烏鰡”又叫青魚,英文叫snakehead )。
但中西有志一同的是:皆找不到鰻種魚或成熟鰻魚(大腹便便,富含魚卵者),所以都瞎掰。但中國人比較相信生源論,除了孫行者外,凡事皆有個源頭。便創出鰻魚騙烏鳢幫他生小孩的傳說 (這大概是最早的surrogate概念),從此以後鰻變成複姓-鰻鱺(鳢),頗有冠夫姓之味道。而台灣通史誌的紀載,大抵可以看出,文抄公引用文獻之痕 (摘自古今圖書集成),但比較特殊的是台灣人管鰻叫烏鰻,這點沒錯;而且殺鰻發現有蛋者,八成有毒,不要吃它,這點值得玩味,直到現在,未有文獻記載大腹便便,富含魚卵者,故人皆曰可殺,對合理吃鰻魚找到歷史文獻的支持。
臨國日本,認為鰻魚是山藥或蕃薯變來的,因為兩者皆為深色長條狀、富含黏液,更重要的是鰻魚可以利用皮膚呼吸,有一點濕潤就可以活,所以看到一條黑不溜丟的鰻在山野小徑散步,不得不連想,它和山藥的血緣關係 (小日本,連想像力都早已 脫亞入歐…)。
Subscribe to:
Comments (Atom)